Milline lõhustumine toimub geenide polümeerse toime käigus. Mittealleelsete geenide koostoime: komplementaarsus, epistaas, polümeer, pleiotroopia. Kromosomaalne pärilikkuse teooria
Alleelsete geenide ja selle tüüpide koostoime
G. Mendeli seadusi uurides oleme juba tutvunud alleelsete geenide põhiliste interaktsiooni tüüpidega. Eelnevalt uuritud materjali põhjal täitke tabel.
Tabel. Ühe alleelse paari geenide interaktsiooni tüübid
Interaktsiooni tüüp |
Interaktsiooni olemus |
Valemid fenotüübiliseks lõhustamiseks F 2-s ja testristamisel |
Näited |
| Täielik domineerimine | Üks alleel (dominantne) pärsib täielikult teise (retsessiivse) alleeli mõju |
Kõrge taimekasvu domineerib kääbus; lokkis juuksed - üle sirgete juuste; pruunid silmad - siniste kohal jne. |
|
| Mittetäielik domineerimine | Iga alleeli mõju ei realiseeru täielikult, mille tulemusena ilmneb vahepealne tunnus |
Öiste ilulillede värvimine; sirprakuline aneemia; tsüstonuuria jne. |
|
| Kaasdominantsus | Mõlema alleeli osalemine tunnuse määramisel |
IV veregrupp (selle veregrupi erütrotsüüdid sisaldavad seda tüüpi aglutinogeene A ja tüüp IN) |
Mittealleelsete geenide interaktsioon
G. Mendeli seaduste põhjal võime jõuda järeldusele, et geeni ja tunnuse vahel on üsna tugev seos, et genotüüp koosneb iseseisvalt toimivate geenide summast ning fenotüüp on mehaaniline kombinatsioon. individuaalsetest omadustest. Otsesed ja ühemõttelised seosed geeni ja tunnuse vahel on aga pigem erand kui reegel. Tunnuse areng organismis on tavaliselt paljude geenide kontrolli all ja tunnus on mittealleelsete geenide koosmõju tulemus.
Lihtsaim mittealleelsete geenide interaktsiooni juhtum on see, kui tunnust kontrollivad kaks alleelide paari. Vaatame mõnda näidet sellisest suhtlusest.
Täiendav geenide interaktsioon
Geenide interaktsiooni nähtust, mille puhul on tunnuse avaldumiseks vajalik kahe mittealleelse geeni olemasolu, nimetatakse täiendavus(alates lat. komplementum- lisamine) ja tunnuse avaldumiseks vajalikud geenid on täiendavad või täiendav.
Vaatleme erinevaid võimalusi täiendavaks geenide interaktsiooniks.
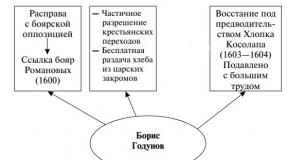
Teise geeni alleelid ilmnevad ainult geeni domineeriva alleeli juuresolekulA. Hiirte värvus sõltub kõige lihtsamal juhul kahest geenist. Genotüübiga hiired ahh puudub pigment ja on valge. Domineeriva alleeli olemasolul A pigment toodetakse ja hiir on kuidagi värviline. Konkreetse värvi määrab teine geen. Selle domineeriv alleel KOOS põhjustab hiire halli värvi ja retsessiivse alleeli Koos- must värv. Seega, kui arvestada homosügootseid variante, on hallide hiirte genotüüp AASS, must - AAAss, valge - aaSS või aass. Halli hiire ületamisel AASS valgega aass esimeses põlvkonnas selgub kõik Mendeli järgi: kõik hübriidid on halli värvi (need on genotüübiga heterosügootid AaSs). F2-s, nagu on lihtne kontrollida, saame 9/16 hallid hiired, 3/16 mustad ja 4/16 valged.

Et tunnus avalduks, peavad genotüübis olema kahe erineva geeni domineerivad alleelid. Sellise komplementaarse geeniinteraktsiooni näide on kõrvitsa viljakuju pärandumine. Mõlema domineeriva alleeli olemasolul on viljad kettakujulised, ühe (ükskõik millise!) domineeriva alleeli olemasolul - sfäärilised, domineerivate alleelide puudumisel - piklikud.

Tunnuse avaldumiseks peavad genotüübis esinema kahe erineva geeni domineerivad alleelid, kuid igal domineerival alleelil on kombinatsioonis teise paari retsessiivsete alleelidega sõltumatu fenotüübiline ilming. Näiteks kanadel määrab hernekujulise harja üks domineeriv geen, roosikujulise hari aga teine mittealleelne, aga ka domineeriv geen. Kui need geenid on samas genotüübis, tekib pähklikujuline hari. Kui organism on mõlema retsessiivse geeni suhtes homosügootne, tekib lihtne lehekujuline kamm. Dihübriidide ristamisel (kõik pähklikujulise kammiga) toimub teises põlvkonnas lõhenemine vahekorras 9:3:3:1. Kuid siin on võimatu leida iga alleeli sõltumatut segregatsiooni suhtega 3: 1, kuna mõlema domineeriva geeni genotüübi kokkulangevuse korral nende otsest mõju ei tuvastata.
Seega räägime mittealleelsete geenide komplementaarsest interaktsioonist juhul, kui kaks mittealleelset geeni annavad uue tunnuse, st tekib uus moodustis.
Epstaatiline geenide interaktsioon
Geenide interaktsiooni, kus üks geen surub alla teise, esimese suhtes mittealleelse geeni toime, nimetatakse epistaas(kreeka keelest epi– üleval ja seisak- seisma) ja kutsutakse geene, mis pärsivad teiste geenide toimet supressorgeenid, või inhibiitorgeenid(alates lat. inhiber– hoidke) või epistaatilised geenid. Supresseeritavat geeni nimetatakse hüpostaatiline.
On domineeriv ja retsessiivne epistaas.
Domineeriv epistaas seotud domineeriva inhibiitorgeeniga. Näiteks geen I määrab kõrvitsa viljade valge värvuse, kui see on olemas, geeni mõju IN ei ilmu. Genotüübiga iiBB või iiВb Kõrvitsa viljad on kollased. Lõpuks, kui mõlemad geenid on esindatud retsessiivsete alleelidega, on kõrvitsa vili roheline. Sama tüüpi saab kasutada koerte, lammaste ja teiste loomade karvkatte värvi pärimiseks.
Retsessiivne epistaas tuvastatakse siis, kui mõne teise geeni retsessiivsed alleelid suruvad alla geeni ekspressiooni. Seda tüüpi epistaasi illustreerib koduhiirte karvkatte värvi pärand. Agouti värvi (punakashall) määrab domineeriv geen A, selle retsessiivne alleel A annab musta värvi. Geen mittealleelsest paarist IN soodustab värvi ja geeni avaldumist b on supressor ja pärsib domineeriva alleeli toimet A ja retsessiivne A. Genotüübiga hiired Аbb fenotüübi poolest eristamatu genotüübiga indiviididest aabb- üleni valge.
Polümeerse geeni interaktsioon
Polümerism– mittealleelsete geenide interaktsiooni tüüp, mille puhul ühe tunnuse kujunemisel mõjutavad mitu paari mittealleelseid geene. Selliseid geene nimetatakse polümeer(kreeka keelest poliitika– palju ja meros- osa). Neid tähistatakse sama tähega, kuid erinevate indeksitega, mis näitavad genotüübi alleelsete paaride arvu, mis määravad konkreetse tunnuse arengu ( A 1 A 1 A 2 A 2).
Polümerisatsioonil on kaks varianti: geenide summeeriva efektiga ja geene üksteisega võimendamata.
Polümeergeenide kumulatiivne toime. Sel juhul sõltub tunnuse avaldumise määr indiviidi genotüübi domineerivate alleelide arvust. Seega määrab nisuterade punase värvuse kaks või enam geenipaari. Nende alleelide kõik domineerivad geenid määravad punase värvi, retsessiivsed geenid aga terade valge värvuse. Üks domineeriv alleel toodab terad, mis ei ole väga värvilised. Kui genotüübis on kaks domineerivat alleeli, suureneb värvi intensiivsus. Ainult siis, kui organism on kõigi retsessiivsete geenide paaride suhtes homosügootne, ei ole terad värvitud. Seega toimub dihübriidide ristamisel lõhenemine vahekorras 15 värvilist ja üks valge. Kuid 15 värvitud värvist on üks intensiivne punane värv, sest... sisaldab nelja domineerivat alleeli, neli on veidi heledama värviga, sest sisaldavad genotüüpides kolme dominantset alleeli ja ühte retsessiivset alleeli, kuus on veelgi heledamad kahe domineeriva ja kaks retsessiivset alleeli genotüüpides, neli on veelgi heledamad, sest neil on ainult üks domineeriv ja kolm retsessiivset geeni, see tähendab, et tegelik jaotus on 1: 4: 6: 4: 1.
Selle polümeeri versiooni kohaselt on nahavärv, pikkus ja kaal inimestel päritud. Sarnane pärandumismehhanism on iseloomulik paljudele kvantitatiivsetele, sealhulgas majanduslikult väärtuslikele tunnustele: suhkrusisaldus peedijuurtes, vitamiinide sisaldus puu- ja juurviljades, teravilja kõrre pikkus, maisikõrva pikkus, loomade viljakus, kariloomade piimatoodang. , kanade munatootmine jne.
Polümeergeenid ei võimenda üksteist. Sel juhul on F 2 jagamine 15:1. Seega võivad karjase rahakoti viljad olla kolmnurksed (domineerivad) ja ovaalsed (retsessiivsed). Seda tunnust kontrollivad kaks paari polümeerseid geene. Kui taime genotüübis on vähemalt üks domineeriv alleel esimesest või teisest polümeerigeenide paarist, on vilja kuju kolmnurkne ( A 1 A 2; A 1 ja 2; a 1 A 2 ). Ainult need taimed, mille genotüübis pole ühtegi domineerivat alleeli, on ovaalse vilja kujuga - a 1 a 1 a 2 a 2.
Seega võib teatud alleelide kuhjumine genotüübis kaasa tuua muutusi tunnuste avaldumises.
Mitme geeni toime
 Sageli mõjutab geen organismis mitte ühte, vaid mitmeid tunnuseid. Nähtust, mille puhul üks geen võib mõjutada organismi mitme tunnuse kujunemist, nimetatakse pleiotroopia(kreeka keelest pleon– arvukamad ja tropos- pööre).
Sageli mõjutab geen organismis mitte ühte, vaid mitmeid tunnuseid. Nähtust, mille puhul üks geen võib mõjutada organismi mitme tunnuse kujunemist, nimetatakse pleiotroopia(kreeka keelest pleon– arvukamad ja tropos- pööre).
Selle nähtuse olemasolu ei ole sugugi vastuolus klassikalise kontseptsiooniga “üks geen – üks valk – üks tunnus”. Lihtsustatult võib ühe geeni mõju mitmele tunnusele korraga kujutada järgmiselt. Geenist info lugemise tulemusena moodustub teatud valk, mis saab seejärel osaleda erinevates organismis toimuvates protsessides, avaldades seeläbi mitmekülgset mõju. Näiteks hernestel sõltuvad ühest geenist seemnekesta pruun värvus ja pigmendi areng taime teistes osades ning õite värvus; Drosophilas mõjutab silmade valget värvi põhjustav geen kehavärvi heledamaks muutumist ja muutusi mõnes siseorganis; Inimestel määrab punaste juuste eest vastutav geen ka heledama nahavärvi ja tedretähnide väljanägemise. Marfani sündroomi põhjustab autosoomne domineeriv pleiotroopne geen ja see väljendub suures kasvus, sõrmede ja varvaste (ämbliksõrmed) luude pikenemises, liigeste hüpermobiilsuses, silmaläätse subluksatsioonis ja südamehaigustes.
See mitmekordne toiming on tüüpiline enamikule geenidele. Siiski ei tohiks ette kujutada, et pleiotroopne geen mõjutab iga tunnust võrdselt. Valdav enamus erineva pleiotroopia astmega geene iseloomustab tugevam mõju ühele tunnusele ja palju nõrgem mõju teisele.
Genotüüp kui terviklik süsteem
Hübriidide lõhenemine järglastes võimaldab meil väita, et genotüüp koosneb üksikutest elementidest - geenidest, mida saab pärida iseseisvalt ( genotüübi diskreetsus). Samas ei saa genotüüpi pidada üksikute geenide lihtsaks mehaaniliseks summaks. Genotüüp on interakteeruvate geenide süsteem. Täpsemalt ei interakteeru mitte geenid ise (DNA lõigud), vaid nende alusel moodustunud geeniproduktid (RNA ja seejärel valgud). Seetõttu on mõnel juhul erinevate geenide toime suhteliselt sõltumatu, kuid reeglina on tunnuse avaldumine erinevate geenide produktide koosmõju tulemus.
Mis tahes organismi genotüüp on interakteeruvate geenide kompleksne terviklik süsteem. See genotüübi terviklikkus tekkis ajalooliselt liigi evolutsiooniprotsessis. Mutatsioonide tulemusena tekivad pidevalt uued geenid, tekivad uued kromosoomid ja isegi uued genoomid. Äsja tekkinud geenid võivad koheselt suhelda olemasolevate geenidega või muuta viimaste töö olemust, isegi olles retsessiivsed, st ilma end avaldamata.
Järelikult avaldub genotüüp igal taime- ja loomaliigil tervikliku süsteemina, mis on antud hetkel ajalooliselt välja kujunenud.
II. Teadmiste kinnistamine
Vestluse kokkuvõtte tegemine uue materjali õppimisel.
III. Kodutöö
Uurige õpiku lõiku (geenide ja tunnuste vaheline seos, alleelsete ja mittealleelsete geenide interaktsiooni tüübid, pleiotroopia, genotüüp kui terviklik süsteem).
Lahenda õpiku lõigu lõpus olevad ülesanded.
Inimese genotüüp sisaldab tohutul hulgal geene, mis kannavad teavet meie keha omaduste ja omaduste kohta. Vaatamata nii suurele arvule toimivad nad ühtse integreeritud süsteemina.
Koolibioloogia kursusest tunneme Mendeli seaduspärasusi, kes uuris tunnuste pärimise mustreid. Uurimistöö käigus avastas teadlane domineerivad ja retsessiivsed geenid. Mõned suudavad teiste ilminguid maha suruda.
Tegelikult ulatub geenide koostoime Mendeli seadustest palju kaugemale, kuigi järgitakse kõiki pärimise reegleid. On võimalik näha erinevusi fenotüübilise segregatsiooni mustris, kuna interaktsiooni tüüp võib erineda.
Geeni omadused
Geen on pärilikkuse ühik, millel on teatud omadused:
- Geen on diskreetne. See määrab konkreetse tunnuse arenguastme, sealhulgas biokeemiliste reaktsioonide omadused.
- Mõjub järk-järgult. Keharakkudesse akumuleerudes võib see põhjustada sümptomi avaldumise suurenemist või vähenemist.
- Kõik geenid on rangelt spetsiifilised, see tähendab, et nad vastutavad konkreetse valgu sünteesi eest.
- Ühel geenil võib olla mitu mõju, mis võib korraga mõjutada mitme tunnuse arengut.
- Ühe tunnuse kujunemises võivad osaleda erinevad geenid.
- Kõik geenid võivad üksteisega suhelda.
- Geeni toime avaldumist mõjutab väliskeskkond.
Geenid on võimelised toimima kahel erineval tasemel. Esimene on geneetiline süsteem ise, milles määratakse geenide seisund ja nende töö, stabiilsus ja varieeruvus. Teise tasemega võib arvestada keharakkudes töötades.
Alleelsete geenide interaktsiooni tüübid
Kõigil meie keharakkudel on diploidne kromosoomide komplekt (nimetatakse ka kahekordseks). Muna 23 kromosoomi ühinevad sama arvu sperma kromosoomidega. See tähendab, et iga tunnust esindab kaks alleeli, seega nimetatakse neid alleeligeenideks.
Sellised alleelipaarid tekivad viljastamise käigus. Need võivad olla kas homosügootsed, st koosneda identsetest alleelidest, või heterosügootsed, kui esinevad erinevad alleelid.
Alleelsete geenide interaktsiooni vormid on selgelt esitatud tabelis.
| Interaktsiooni tüüp | Interaktsiooni olemus | Näide |
| Täielik domineerimine | Domineeriv geen surub täielikult alla retsessiivse geeni avaldumise. | Hernevärvi, inimese silmavärvi pärand. |
| Mittetäielik domineerimine | Domineeriv geen ei suru täielikult alla retsessiivse geeni ekspressiooni. | Öise ilu (lill) lillevärvimine. |
| Kaasdominantsus | Heterosügootses olekus põhjustab iga alleelgeen oma kontrollitava tunnuse arengut. | Veregrupi pärand inimestel. |
| Üledomineerimine | Heterosügootses seisundis ilmnevad sümptomid selgemalt kui homosügootses seisundis. | Ilmekas näide on heteroosi nähtus loomadel ja taimestik, sirprakuline aneemia inimestel. |
Täielik ja mittetäielik domineerimine
Täielikust domineerimisest saame rääkida juhul, kui üks geenidest suudab tagada tunnuse avaldumise, kuid teine ei suuda seda teha. Tugevat geeni nimetatakse domineerivaks ja selle vastast retsessiivseks.
Pärand toimub sel juhul täielikult Mendeli seaduste järgi. Näiteks herneseemnete värvus: esimeses põlvkonnas näeme, et kõik herned on rohelised, see tähendab, et see värv on domineeriv omadus.
Kui viljastamise käigus saavad kokku pruunide silmade ja siniste silmade geen, siis on lapsel pruunid silmad, sest see alleel pärsib täielikult siniste silmade eest vastutava geeni.

Mittetäieliku domineerimise korral võib heterosügootides näha vahepealse tunnuse avaldumist. Näiteks ristades punaste õitega homosügootset öökaunitari domineeriva tunnuse saamiseks sama isendiga, ainult valge korollaga, võib esimeses põlvkonnas näha roosasid hübriide. Domineeriv punane tunnus ei suru täielikult alla retsessiivse valge tunnuse avaldumist, nii et lõpuks saame midagi vahepealset.
Kaasdominantsus ja üledominantsus
Seda geenide koostoimet, milles igaüks annab oma tunnuse, nimetatakse kodominantsiks. Kõik geenid ühes alleelpaaris on absoluutselt samaväärsed. Kumbki ei saa teise tegevust maha suruda. Just seda geenide koostoimet täheldame inimestel veregruppide pärimisel.
Geen O tagab 1. veregrupi avaldumise, geen A - teine, geen B - kolmas ja kui geenid A ja B leitakse koos, siis kumbki ei suuda teise avaldumist maha suruda, nii moodustub uus tunnus - veregrupp 4.
Üledominantsus on veel üks näide alleelsete geenide koostoimest. Sel juhul heterosügootsed isikud see omadus neil on homosügootsetega võrreldes rohkem väljendunud ilming. See geenide interaktsioon on sellise nähtuse nagu heteroos (hübriidse elujõu nähtus) aluseks.

Näiteks kahe tomatisordi ristamise korral saadakse hübriid, mis pärib mõlema algse organismi omadused, kuna omadused muutuvad heterosügootseks. Järgmises põlvkonnas algab lõhenemine omaduste järgi, nii et samu järglasi pole võimalik saada.
Loomamaailmas võib isegi jälgida selliste hübriidvormide viljatust. Selliseid geenide interaktsiooni näiteid võib leida sageli. Näiteks eesli ja mära ristamise korral sünnib muul. Ta päris kõik parimad omadused tema vanemad, kuid ta ise ei saa järglasi.
Inimestel on sirprakuline aneemia selle tüübi järgi päritav.
Mittealleelsed geenid ja nende koostoime
Geene, mis paiknevad erinevates kromosoomipaarides, nimetatakse mittealleelseteks. Kui nad leiavad end koos, võivad nad üksteist hästi mõjutada.
Mittealleelsete geenide interaktsioon võib toimuda mitmel viisil:
- Vastastikune täiendavus.
- Epistaas.
- Polümeeri toime.
- Pleiotroopia.
Kõigil seda tüüpi geenide interaktsioonidel on oma eripärad.
Vastastikune täiendavus
Selle interaktsiooniga täiendab üks domineeriv geen teist, mis on samuti domineeriv, kuid ei ole alleelne. Kui nad kokku tulevad, aitavad nad kaasa täiesti uue iseloomuomaduse avaldumisele.

Võite tuua näite magusate herneste lillede värvi ilmnemisest. Pigmendi ja seega ka värvi olemasolu lilles tagab kahe geeni – A ja B – kombinatsioon. Kui vähemalt üks neist puudub, on õis valge.
Inimestel täheldatakse sellist mittealleelsete geenide koostoimet kuulmisorgani moodustumise ajal. Normaalne kuulmine saab tekkida ainult siis, kui mõlemad geenid – D ja E – on domineerivas olekus. Kui retsessiivses seisundis on ainult üks dominant või mõlemad, pole kuulmist.
Epistaas
See mittealleelsete geenide interaktsioon on täiesti vastupidine eelmisele interaktsioonile. Sel juhul on üks mittealleelne geen võimeline teise ekspressiooni alla suruma.
Selle variandi geenide interaktsiooni vormid võivad olla erinevad:
- Domineeriv epistaas.
- Retsessiivne.
Esimest tüüpi interaktsiooni korral pärsib üks domineeriv geen teise domineeriva geeni ekspressiooni. Retsessiivsed geenid on seotud retsessiivse epistaasiga.

Seda tüüpi koostoime põhjustab kõrvitsatel viljavärvi ja hobustel karvavärvi pärilikkust.
Geenide polümeerne toime
Seda nähtust võib täheldada, kui sama tunnuse avaldumise eest vastutavad mitu domineerivat geeni. Kui esineb vähemalt üks domineeriv alleel, ilmneb see tunnus kindlasti.
Geeni interaktsioonide tüübid võivad sel juhul olla erinevad. Üks neist on akumulatiivne polümerism, kui tunnuse avaldumisaste sõltub domineerivate alleelide arvust. Nii pärandub nisuterade värvus ehk inimese nahavärv.

Kõik teavad, et kõigil inimestel on erinev nahavärv. Mõnel on nahk täiesti hele, mõnel tume nahk ja negroidide rassi esindajatel on nahk täiesti must. Teadlased on seisukohal, et nahavärvi määrab kolme erineva geeni olemasolu. Näiteks kui kõik kolm esinevad genotüübis domineerivas olekus, siis on nahk kõige tumedam, nagu mustanahalistel.
Otsustades meie nahavärvi järgi, ei ole kaukaasia rassil domineerivaid alleele.
Ammu on välja selgitatud, et mittealleelsete geenide interaktsioon vastavalt polümeeri tüübile mõjutab enamikku inimese kvantitatiivseid tunnuseid. Nende hulka kuuluvad: pikkus, kehakaal, intellektuaalsed võimed, keha vastupidavus nakkushaigused ja mõned teised.
Võib vaid märkida, et selliste märkide areng sõltub keskkonnatingimustest. Inimesel võib olla eelsoodumus ülekaalule, kuid dieeti järgides on võimalik seda probleemi vältida.
Geenide pleiotroopne toime
Teadlased on juba ammu veendunud, et geenide interaktsioonide tüübid on üsna mitmetähenduslikud ja väga mitmekülgsed. Mõnikord on teatud fenotüübiliste tunnuste avaldumist võimatu ennustada, sest pole teada, kuidas geenid omavahel suhtlevad.
Seda väidet rõhutab vaid nähtus, et üks geen võib mõjutada mitme tunnuse teket ehk omada pleiotroopset toimet.
Pikka aega on täheldatud, et punase pigmendi olemasoluga peedi viljades kaasneb tingimata sama, kuid ainult lehtede olemasolu.
Inimestel on teadaolev haigus, mida nimetatakse Marfani sündroomiks. Seda seostatakse arengu eest vastutava geeni defektiga sidekude. Selle tulemusena selgub, et kõikjal, kus see kude kehas asub, võib probleeme täheldada.

Sellistel patsientidel on pikad "ämbliku" sõrmed ja neil on diagnoositud silmalääts või südamerike.
Keskkonnategurite mõju geenide tegevusele
Ei saa eitada väliskeskkonna tegurite mõju organismide arengule. Nende hulka kuuluvad:
- Toitumine.
- Temperatuur.
- Valgus.
- Pinnase keemiline koostis.
- Niiskus jne.
Keskkonnategurid on valiku, pärilikkuse ja varieeruvuse protsessides põhilised.
Kui käsitleme alleelsete või mittealleelsete geenide interaktsiooni vorme, peame alati arvestama keskkonna mõjuga. Võime tuua järgmise näite: kui priimula taimi ristatakse temperatuuril 15-20 kraadi, siis on kõik esimese põlvkonna hübriidid roosat värvi. Temperatuuril 35 kraadi muutuvad kõik taimed valgeks. Niipalju siis keskkonnategurite mõjust tunnuste avaldumisele, siin pole enam tähtsust, milline geen on domineeriv. Küülikutel, selgub, oleneb ka nende karva värvus temperatuuritegurist.
Teadlased on pikka aega tegelenud küsimusega, kuidas kontrollida sümptomite ilminguid erinevate välismõjude abil. See võib anda võimaluse arengut kontrollida kaasasündinud omadused, mis on inimestele eriti oluline. Miks mitte kasutada oma teadmisi mõne päriliku haiguse ilmnemise ennetamiseks?
Kõik alleelgeenide ja mitte ainult nende interaktsiooni tüübid võivad olla nii erinevad ja mitmetahulised, et neid on võimatu ühelegi konkreetsele tüübile omistada. Võib vaid väita, et kõik need vastasmõjud on võrdselt keerulised nii inimestel kui ka kõigi taime- ja loomaliikide esindajatel.
Taas on paslik rõhutada, et tunnuste pärimise mõistet kasutatakse enamasti kujundliku väljendina. Tegelikkuses ei päri mitte tunnused, vaid erinevatest geenidest koosnevad DNA molekulid .
Geenid kodeerivad infot teatud organismi valkude kohta ja need teatud valgud, sünteesitud rakkudes, on juba üks või teine organismile iseloomulik tunnus.
Tunnused ise kujunevad välja organismi individuaalse arengu käigus. Neid määravad ainult need geenid kogu tohutust geenide arsenalist, mida nimetatakse organismi genotüübiks ja mis avalduvad konkreetsetes keskkonnatingimustes. Seetõttu ütleme, et selle määravad genotüüp ja keskkonnategurid.
See artikkel on jätk kahele eelmisele ja käsitleb geneetika küsimust. Nagu varem märgitud, võib geenidel olla kaks erinevat olemust vastavalt nende võimele realiseerida spetsiifilisi omadusi. See on interaktsioonide rakendamine : 1) alleelsed ja 2) mittealleelsed geenid.
Alleelsete geenide interaktsioonide tüüpe käsitletakse artiklis. Selles artiklis kirjeldame mittealleelsete geenide nelja peamist interaktsiooni tüüpi: komplementaarsus, epistaas, polümerisatsioon ja geenide pleiotroopne toime.
Kõigil, kes soovivad mõista mittealleelsete geenide interaktsiooni teemat, soovitan teil osta minu kollektsiooni (kogu sisaldab ka teoreetilise osa ja probleemide lahendusi käsitletakse üksikasjalikult).
1. Täiendav geenide interaktsioon
See on siis, kui üks domineeriv geen täiendab teise mittealleelse domineeriva geeni toimet. Selle tulemusena tekib moodustumine tunnuse uus AVALDUS vastastikuse abi liigi järgi üksteist täiendavad.
Jällegi juhin teie tähelepanu asjaolule, et see pole "uus märk", vaid ainult " sama tunnuse uus ilming" kuna kõigi siin vaadeldavate alleelsete ja mittealleelsete geenide interaktsiooni tüüpide puhul räägime alati ühe tunnuse (kuid selle avaldumise erinevatest vormidest) pärilikkuse uurimisest.
Ärge olge segaduses täiendavad interaktsiooni mittealleelne geenid (st täiesti erinevad geenid, kuid vastutavad ka ühe tunnuse avaldumise eest) koos nähtusega, mis on omane alleelne geenid.
Allolev tabel illustreerib fenotüübi ja genotüübi järgi jagunemise võimalikke juhtumeid F 2 juures täiendavad mittealleelsete geenide interaktsioon. Pange tähele, et sisse F 2 st "diheterosügootide" üksteisega ristumisest.
Miks ma kirjutan jutumärkidesse "diheterosügoot"? Sest need pole üldse Mendeli diheterohüsoodid (st heterosügootsed organismid kahes paaris erinevad märgid ) ja "diheterosügootid", kuigi ka kahe erineva geenipaari jaoks, kuid vastutavad avaldumise eest üks märk.
Magusherne lillede värvipärimise näitel
variandi järgi on võimalik selgelt jälgida geenide komplementaarse interaktsiooni nähtust 2. tabelis loetletud.
Selle taime kahe erineva joone ületamiselvalgelilled F-hübriidides 1 lilled osutusidlilla.
Seega, kui meile pakutud ülesandes näeme, et jagamine fenotüübi järgi on üks neljast ülalolevas tabelis loetletud, siis tähendab see, et selline ülesanne tuleb lahendada, võttes arvesse epistaatiline interaktsioon mittealleelsed geenid.
Tabelist on selge, et valemid 3. Ja 4. on fenotüübilt identsed, kuid genotüübilt erinevad, kuna liinis 4. registreeritakse genotüüpide pärand retsessiivne epistaas ja ridades 1. , 2. Ja 3. - kell domineeriv.
3. Polümeeria
See on siis, kui sama tunnuse avaldumise eest vastutavad mitu erinevat domineerivat geeni.
See tähendab, et geenid on erinevad, kuid vastutavad sama tunnuse avaldumise eest. Nende erinevate geenide vähemalt ühe domineeriva alleeli olemasolu genotüübis põhjustab domineeriva tunnuse väljakujunemise ja retsessiivne tunnus areneb ainult siis, kui genotüübis on ainult retsessiivsed alleelid.
Tunnuse avaldumise intensiivsus sõltub domineerivate geenide arvust genotüübis. Tavaliselt määravad need geenid kvantitatiivsed tunnused (kasvu intensiivsus, munatoodang, rasvasisaldus ja piima kogus, vitamiinide sisaldus puuviljades, inimese naha pigmentatsiooni intensiivsus, kõrvade või teravilja terade värvuse intensiivsus jne). Kuid saab määrata ka kvalitatiivseid märke.
Polümeergeenid on tähistatud ühe tähega (mitte segi ajada ühe tunnuse alleelsete geenidega!), kuid erineva numeratsiooniga, et näidata, et tegemist on erinevate geenidega, nt. A 1, A 2, A 3 jne.
Polümeeride koostoimet ja geenide toimet käsitletakse tavaliselt erinevate nisusortide ristamise näitel valge (a1a1a2a2) Ja tumepunane (A 1 A 1 A 2 A 2) maisikõrvad. Nagu näeme, vastutavad punase värvi intensiivsuse eest kaks erinevat domineerivat geeni. A 1 Ja A 2, ja punase värvuse intensiivsus sõltub sellest, kas organismi genotüübis on iga domineeriva geeni kõik alleelid.
____________________________
P.S. Nüüd oleme lõpetanud tutvumise sissejuhatava osa erinevat tüüpi geenide interaktsioonid. "Sissejuhatav" mõtlen võimaluse eest hakata teadlikult lahendama geneetilisi probleeme. Kuna geneetika probleemide lahendamiseks on oluline teada, et geenide koostoimeid on ainult kahte tüüpi : alleelsete ja mittealleelsete geenide jaoks. Kuid igal neist tüüpidest on oma vormid (peamised neist oleme arutanud).
Teadusliku kogemuse kogunedes ilmnesid vastuolud Mendeli iseseisva pärimise kolmanda seadusega. Järglased jagati fenotüübi järgi suhtega 15:1 või 9:7, mitte Mendeli 9:16. See näitab teatud seoseid mittealleelsete geenide vahel.
mehhanism
Mittealleelsed geenid paiknevad kromosoomide erinevates osades ja kodeerivad erinevat tüüpi valgud. Geenid ei mõjuta üksteist otseselt, mistõttu interaktsioon toimub tsütoplasmas teatud geenide poolt kodeeritud valkude tasemel.

Riis. 1. Mittealleelsed geenid.
Koostoimemehhanism võib toimuda vastavalt ühele kolmest stsenaariumist:
- kahe ensüümi samaaegne toime, mis kodeerivad kahte mittealleelset geeni;
- üks mittealleelne geen toodab valku, mis mõjutab teise mittealleelse geeni tööd (supresseerib või aktiveerib);
- kaks valku, mida kodeerivad kaks mittealleelset geeni, toimivad samal protsessil, tugevdades või taastades sama tunnust.
Üks geen võib vastutada mitme fenotüübilise tunnuse eest või mitu geeni võib olla vastutav ühe tunnuse eest.
Liigid
Mittealleelsete geenide interaktsiooni on mitut tüüpi, peamised neist on üksikasjalikult kirjeldatud tabelis.

Riis. 2. Vastastikune täiendavus.
|
Vaade |
Kirjeldus |
Näide |
|
Kahe erineva geeni põhjustatud tunnus ilmneb ainult kahe domineeriva alleeli kombineerimisel. Selliseid geene nimetatakse komplementaarseteks. Tunnus ei moodustu ühe geeni puudumisel. F2-s esinevate fenotüüpsete märkide lõhenemine toimub vahekorras 9:7, 9:6:1, 9:3:4 |
Magusate herneste ristamine valgete õitega. F1-s on kõigil järglastel lillad õied, sest domineerivate geenide A ja B kombinatsioon kodeerib antotsüaniini, mis annab lilla värvuse. Üksikud geenid ei moodusta lillat. F2-s toimub lõhenemine - 9 lillat (AB), 7 valget (3 - Abb, 3 - aaB, 1 - aabb) |
|
|
Üks geenipaar pärsib teist, takistades fenotüübilise tunnuse ilmnemist. Supresseerivat geeni nimetatakse epistaatiliseks (supressorgeen või inhibiitor), allasurutud geeni nimetatakse hüpostaatiliseks. Inhibiitorit tähistatakse tähega I, st. Epistaas võib olla domineeriv – supressioon domineeriva geeni poolt (I>B, b) ja retsessiivne – supressioon retsessiivse geeni poolt (i>B,b). Domineeriva seisundi korral toimub geenide lõhenemine vahekorras 7:6:3, 12:3:1, 13:3, retsessiivse manifestatsiooniga - 9:3:4, 9:7, 13:3 |
Kaera tera värvus: A - must, B - hall. F1-s on kõik terad mustad, kui geen A on epistaatiline (AaBB või IiBB). F2-s toimub jagamine vastavalt tera värvile - 12 musta, 3 halli ja 1 valget. 12 taimes on I geen tingimata olemas, 3 taimes on see retsessiivses olekus - st. Üks taim saab iibb geenid (mustade ja hallide värvide puudumine), seega on see valge |
|
|
Polümerism |
Kvantitatiivsed või dimensioonilised tunnused, mida ei saa fenotüübi järgi selgelt jaotada (pikkus, piimakogus, kariloomade rasvasisaldus), määrab geenide kogum. On kumulatiivseid ja mittekumulatiivseid tüüpe. Esimesel juhul sõltub tunnuse avaldumine geenide toimete summast (mida domineerivamad geenid, seda heledam on tunnus). Teisel juhul avaldub tunnus domineeriva geeniga, geenide arv ei mõjuta fenotüübi avaldumist. Kumulatiivsel kujul F2-s täheldatakse lõhenemist vahekorras 1:4:6:4:1, mittekumulatiivsel kujul - 15:1. Polümeergeenid on tähistatud ühe tähega (A, a, B, b jne) ja alleelid on tähistatud numbriga. Näiteks A1a1A2a2 |
Inimese nahavärv sõltub nelja geeni toimest: A1A1A2A2 - must, a1a1a2a2 - valge, A1A1A2a2, A1a1A2A2, A1a1A2a2, A1A1a2a2, a1a1A2A2, A1a1a2a2, a1a1 medium toon (mustast kuni mossalt) |

Riis. 3. Epistaas.
Geenide mitut toimet nimetatakse pleiotroopiaks. Ühe geeni toime on tavaliselt tingitud interaktsioonist teiste geenidega. Enamikul geenidel on selline toime, seega on genotüüp interakteeruvate geenide süsteem.
TOP 4 artiklitkes sellega kaasa loevad
Mida me õppisime?
Õppisime lühidalt mittealleelsete geenide interaktsiooni tüüpidest. Interaktsioone on kolme tüüpi – komplementaarsus, epistaas, polümerisatsioon. Tunnuse komplementaarseks avaldumiseks on vajalik kahe domineeriva geeni olemasolu. Epistaasi iseloomustab teise geeni toime pärssimine ühe geeni poolt. Polümerism on geenide komplekti koostoime. Paljude geenide koostoimet nimetatakse pleiotroopiaks.
Test teemal
Aruande hindamine
Keskmine hinnang: 4.6. Kokku saadud hinnanguid: 90.
Alleelsete geenide interaktsioon esineb kolmel kujul: täielik domineerimine, mittetäielik domineerimine ja sõltumatu manifestatsioon (kodominantsus).
Täielik domineerimine - kui üks domineeriv alleel surub täielikult alla retsessiivse alleeli avaldumise, näiteks on herneste kollane värvus domineeriv rohelise värvi üle.
Mittetäielikku domineerimist täheldatakse siis, kui alleelipaarist üks geen ei taga oma valguprodukti moodustumist, mis on piisava tunnuse normaalseks avaldumiseks. Selle geenide interaktsiooni vormi korral erinevad kõik heterosügootid ja homosügootid üksteisest fenotüübi poolest oluliselt. Mittetäieliku domineerimisega lõhenemise näide on Night Beauty lillede värvi pärand.
Punaste õitega (AA) ja valgete õitega (aa) taimede ristamisel on F1 hübriididel roosad õied (Aa). Seega tekib mittetäielik domineerimine; F2-s on nii fenotüübi kui ka genotüübi jagunemine 1:2:1.
Lisaks täielikule ja mittetäielikule domineerimisele on teada domineerivate-retsessiivsete suhete või kaasdominantsi puudumise juhtumeid. Kui heterosügootsetes organismides kodominantne, põhjustab iga alleelgeen fenotüübis tema poolt kontrollitava tunnuse moodustumist.
Sellise alleelide interaktsiooni vormi näide on inimese veregruppide pärand vastavalt AB0 süsteemile, mis on määratud I geeniga Selle geeni Io, Ia, Ib alleeli määravad veregrupi antigeenid. Veregruppide pärandumine illustreerib ka hulgialleelismi fenomeni: inimpopulatsioonide genofondides eksisteerib geen I kolme erineva alleeli kujul, mis on üksikutel indiviididel kombineeritud vaid paarikaupa. Enne seda näidet rääkisime geenidest, mis eksisteerivad ainult kahes erinevas alleelses vormis. Paljud geenid koosnevad aga sadadest nukleotiidipaaridest, mistõttu mutatsioonid võivad esineda paljudes geenipiirkondades ja tekitada palju erinevaid alleelvorme. Kuna igal homoloogsel kromosoomil on üks alleelne geen, siis loomulikult ei ole diploidsel organismil populatsiooni genofondis rohkem kui kaks alleeli seeriat.
30. Mittealleelsed geenide interaktsioonid
Mittealleelsed geenid- need on geenid, mis asuvad kromosoomide erinevates osades ja kodeerivad erinevaid valke. Mittealleelsed geenid võivad ka omavahel suhelda.
Sel juhul määrab kas üks geen mitme tunnuse väljakujunemise või vastupidi, üks tunnus avaldub mitme geeni kombinatsiooni mõjul. Mittealleelsetel geenidel on kolm vormi ja interaktsiooni:
täiendavus;
polümerism.
Täiendav(täiendav) geenitegevus on mittealleelsete geenide interaktsiooni tüüp, mille domineerivad alleelid genotüübis kombineerituna määravad tunnuste uue fenotüübilise ilmingu. Sel juhul võib F2 hübriidide jagunemine fenotüübi järgi toimuda vahekordades 9:6:1, 9:3:4, 9:7, mõnikord 9:3:3:1. Täiendavuse näide on kõrvitsa vilja kuju pärilikkus. Domineerivate geenide A või B olemasolu genotüübis määrab vilja sfäärilise kuju ja retsessiivsete geenide pikliku kuju. Kui genotüüp sisaldab nii domineerivaid geene A kui ka B, on vilja kuju diskoidne. Puhaste joonte ületamisel sfäärilise viljakujuga sortidega, esimene hübriidpõlvkond F1 kõik viljad on kettakujulised ja F2 põlvkonnas toimub fenotüübi lõhenemine: igast 16 taimest on 9 viljad kettakujulised, 6 - kerajad ja 1 - piklikud.
Epistaas- mittealleelsete geenide interaktsioon, kus üks neist on teise poolt alla surutud. Supresseerivat geeni nimetatakse epistaatiliseks, allasurutud geeniks hüpostaatiliseks. Kui epistaatilisel geenil ei ole oma fenotüübilist ilmingut, siis nimetatakse seda inhibiitoriks ja tähistatakse tähega I. Mittealleelsete geenide epistaatiline interaktsioon võib olla domineeriv ja retsessiivne. Domineeriva epistaasi korral pärsib hüpostaatilise geeni (B, b) avaldumist domineeriv epistaatiline geen (I > B, b). Fenotüübi lõhustumine domineeriva epistaasi ajal võib toimuda vahekorras 12:3:1, 13:3, 7:6:3. Retsessiivne epistaas on hüpostaatilise geeni (i > B, b) alleelide allasurumine epistaatilise geeni retsessiivse alleeli poolt. Fenotüübiline lõhustumine võib toimuda vahekorras 9:3:4, 9:7, 13:3.
Polümerism- mittealleelse mitme geeni interaktsioon, mis selgelt mõjutavad sama tunnuse arengut; Tunnuse avaldumise määr sõltub geenide arvust. Polümeergeenid on tähistatud samade tähtedega ja sama lookuse alleelidel on sama indeks.
Mittealleelsete geenide polümeerne interaktsioon võib olla kumulatiivne ja mittekumulatiivne. Kumulatiivse (akumulatiivse) polümerisatsiooni korral sõltub tunnuse avaldumise määr geenide kokkuvõtvast toimest. Mida domineerivamad geenialleelid, seda rohkem väljendub konkreetne tunnusjoon. F2 lõhustumine fenotüübis toimub vahekorras 1:4:6:4:1.
Mittekumulatiivse polümerisatsiooni korral ilmneb see tunnus vähemalt ühe polümeeri geenide domineeriva alleeli juuresolekul. Dominantsete alleelide arv ei mõjuta tunnuse ekspressiooniastet. Fenotüüpne segregatsioon toimub vahekorras 15:1.
Näide: inimese nahavärv, mis sõltub neljast geenist.