¿Dónde se encuentra el tejido conectivo? Funciones del tejido conectivo. El propio tejido conectivo
Aunque los tejidos conectivos y de sostén parecen diferentes, están estrechamente relacionados porque tienen un origen común. Ambos tejidos se originan a partir del mesénquima: tejido conectivo embrionario. Los tejidos conectivos y de soporte incluyen sustancia tanto celular como intercelular (matriz extracelular, sustancia fundamental). La sustancia intercelular puede ser líquida o más o menos sólida. Ambos tipos de tejido forman estructuras conectivas y de soporte, pero de forma cualitativa y cuantitativamente diferentes. Cuanto menos desempeñan una función de apoyo, más evidente se vuelve su participación en los procesos metabólicos, ya que el tejido conectivo está en contacto con la sangre. Como su nombre indica, este tipo de tejido conecta los órganos con los vasos sanguíneos, aunque también cumple otras funciones. El tejido de soporte incluye tejido conectivo denso, así como hueso y cartílago, que realizan principalmente una función de soporte. Los huesos están bien abastecidos de sangre.
FUNCIONES
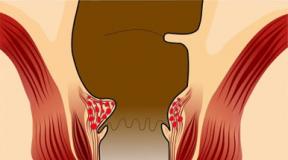
- Función de conexión. Normalmente, el tejido conectivo forma cápsulas de órganos, así como vainas nerviosas y membranas vasculares, y conecta órganos entre sí. En forma de ligamentos, sostiene las articulaciones y en forma de tendones, transmite fuerzas del músculo al hueso.
- Función de intercambio. Aunque los procesos metabólicos ocurren en los fibroblastos, el intercambio de metabolitos ocurre en el ambiente intercelular. Los nutrientes contenidos en la sangre se difunden hacia el entorno intercelular. Desde allí ingresan a las células. Por tanto, el tejido conectivo realiza una función trófica. En consecuencia, las sustancias que salen de las células con la participación del tejido conectivo ingresan a los capilares y vasos linfáticos.
- Balance de agua. La mayor parte del líquido extracelular se encuentra en el espacio intercelular del tejido conectivo areolar (laxo), en el que puede
Se concentrará una gran cantidad de agua. En casos de enfermedades cardíacas y renales, el exceso de líquido en los tejidos puede provocar hinchazón.
- Cicatrización de la herida. Las heridas cicatrizan mediante la formación de tejido conectivo (tejido de granulación), seguido de su endurecimiento y formación de una cicatriz.
- Proteccion. Algunas células especializadas del tejido conectivo que se encuentran en un “estado libre” ( Varios tipos leucocitos), protegen el cuerpo de microbios patógenos y sustancias extrañas. Tienen la capacidad de fagocitar (capturar partículas) y apoyar las funciones protectoras del cuerpo mediante la formación de anticuerpos.
- Funciones tróficas. El tejido adiposo (adiposo) sirve como reserva nutricional del cuerpo.
CÉLULAS DEL TEJIDO CONECTIVO
Entre las células ubicadas en el espacio que ocupa el tejido conectivo, se encuentran los fibroblastos que tienen especificidad tisular. Estas células a veces se denominan fibrocitos, especialmente si están inactivas. Los fibroblastos producen componentes de la sustancia intercelular (sustancia fundamental y estructuras fibrosas). Otro tipo de células que se encuentran son las células que han dejado sistema vascular y convertirse en parte del sistema inmunológico del cuerpo. Estas son las "células sueltas" del tejido conectivo. Son capaces de realizar movimientos ameboides. Según los conceptos modernos, las células libres se originan a partir del mesénquima embrionario y casi todas pertenecen a los glóbulos blancos (leucocitos), que migraron al tejido conectivo desde la sangre.
MATRIZ INTERCELULAR (SUSTANCIA BÁSICA)
Dado que la sustancia intercelular está representada por dos componentes, el tejido conectivo funciona como intermediario entre los vasos sanguíneos y los órganos (sustancia fundamental) y como eslabón conector del cuerpo (estructuras fibrosas). La sustancia fundamental está formada por líquido intersticial, proteínas, polisacáridos y glicoproteínas. Las proteínas y los polisacáridos determinan la consistencia del líquido intersticial. Gracias a su capacidad para retener agua, garantizan, por ejemplo, las propiedades elásticas del cartílago articular y la transparencia de la córnea. Las glicoproteínas son parte del glicocálix, ubicadas en las membranas externas de las células y también son componentes de la membrana basal. En parte, realizan funciones mecánicas (participan en la unión de las células a la matriz extracelular) y, aparentemente, también crean una barrera que regula el intercambio de metabolitos entre el espacio intersticial y las células adyacentes.
Las estructuras fibrosas se dividen en tres tipos: colágenas, elásticas y reticulares. Las fibras de colágeno no se estiran y aparecen en lugares donde se desarrolla la tensión (tendones, ligamentos). Las fibras reticulares son flexibles y su extensa red forma la estructura básica de órganos como los ganglios linfáticos y el bazo. Las fibras elásticas son capaces de estirarse fuerte y reversible. Al mismo tiempo, su longitud puede aumentar más de 1,5 veces (vasos sanguíneos).
Tejido areolar (intersticial) laxo
Tejido conectivo areolar (intersticial) laxo forma un estroma que conecta los tejidos individuales de los órganos; también fija nervios y vasos sanguíneos en sus lugares, formando cajas a su alrededor. Este tejido sirve como depósito de agua y permite que otros tejidos se muevan.
Tejido conectivo fibroso denso y blanco.
Tejido conectivo fibroso denso y blanco. Está formado por fibras y un pequeño número de células. Hay dos tipos de tejido: tejido fibroso blanco denso, sin forma y decorado. En el tejido no formado, las fibras de colágeno se ubican en haces entrelazados entre sí (cápsulas de órganos, capa reticular de la dermis, esclerótica, duramadre del cerebro). En el tejido formado, las fibras de colágeno participan en procesos motores (transferencia de fuerza de los músculos a los huesos). Por tanto, están dispuestos en haces paralelos, visibles a simple vista (por ejemplo, tendones y aponeurosis).
Tejido conectivo reticular
Tejido conectivo reticular muy similar al tejido conectivo embrionario: mesénquima. Se compone de fibras especiales, células reticulares y una extensa red de fibras reticulares. Junto con otros elementos estructurales, el tejido conectivo reticular sirve como marco para los órganos linfáticos (bazo y ganglios linfáticos), cuyos espacios están llenos de "células libres" (por ejemplo, células del sistema inmunológico, linfocitos). En la médula ósea, en el espacio entre las fibras reticulares se encuentran células hematopoyéticas, por lo que el tejido conectivo reticular y las "células libres" constituyen un todo funcional. Al mismo tiempo, las fibras reticulares también se encuentran en el tejido areolar y en los órganos internos (hígado, riñones), donde no forman parte del tejido conectivo reticular. Por ejemplo, las fibras reticulares forman una vaina alrededor de las fibras musculares lisas y estriadas y las unen formando estructuras ordenadas.
Tejido adiposo (adiposo)
Tejido adiposo Es una forma especial de tejido conectivo reticular. Las células del tejido adiposo (lipocitos, adipocitos) acumulan grasa, que se elimina de la sangre mediante el mecanismo de pinocitosis o se forma en las propias células a partir de carbohidratos (azúcares). La gota de grasa ubicada en el adipocito empuja el núcleo celular aplanado hacia la periferia. A lo largo del borde de la célula hay un fino borde de citoplasma. El tejido adiposo realiza funciones mecánicas, es fuente de energía y protege al organismo del frío.
Reserva tejido adiposo. Las grasas sirven como una rica fuente de energía para el cuerpo. Su contenido calórico es el doble que el de los carbohidratos y las proteínas. El tejido conectivo areolar, que forma las vainas de los vasos sanguíneos en el tejido conectivo subcutáneo, sirve como área de almacenamiento para el exceso de grasa. Si es necesario, esta grasa se puede utilizar para las necesidades energéticas del organismo. Al mismo tiempo, las células siguen siendo viables y continúan desempeñando sus funciones de reserva. Según el punto de vista moderno, las células grasas formadas en la primera infancia continúan existiendo durante el resto de la vida de una persona y desempeñan la función de deposición.
Tejido adiposo estructural. A diferencia de la reserva, el tejido adiposo estructural sirve para mantener la forma de partes individuales del cuerpo (plantas de los pies, palmas de las manos, nalgas, mejillas y cuencas de los ojos). Comienza a usarse como reserva de energía solo cuando el cuerpo está muy hambriento (ojos hundidos, mejillas hundidas).
Tejido adiposo marrón. Este tipo de tejido adiposo (tejido adiposo marrón, tejido multilocular) es un tipo especial de tejido adiposo que contiene numerosas mitocondrias oscuras ricas en citocromo. En los recién nacidos se sitúa entre los omóplatos. En los primeros meses de vida, el tejido adiposo pardo cumple una importante función como reservorio de calor. En los adultos, está presente en casos raros, pero es típico de los roedores (proporciona el calentamiento del cuerpo después de la hibernación).
Tejido de soporte
Los tejidos de soporte incluyen tejido óseo y cartilaginoso. Esto también debe incluir tejido cordal y esmalte dental, tejido óseo especializado que es muy duradero. Estos tejidos están compuestos principalmente por fibras de colágeno, lo que confiere rigidez a su estructura. La resistencia del cartílago a las cargas mecánicas está garantizada por la estructura especial de la matriz extracelular, y la resistencia del hueso está asociada con el depósito de sales de calcio en él.
Tejido de cuerda
tejido de cuerda su estructura se asemeja al tejido adiposo, con la única excepción de que en lugar de grasa, las células contienen líquido. Este tejido se encuentra en los vertebrados, incluidos los humanos, donde está representado por el órgano embrionario primario, la notocorda (chorda dorsalis; cuerda dorsal). Debido al denso empaquetado de células, la notocorda es duradera y elástica, como el neumático de la rueda de un automóvil. En un adulto, la notocorda se redujo, quedando solo en la forma del núcleo pulposo de los discos intervertebrales (núcleo pulposo).
tejido cartilaginoso
tejido cartilaginoso Localizado en el esqueleto y el tracto respiratorio. Las características de este tejido son las células cartilaginosas (condrocitos). Se encuentran en la sustancia cartilaginosa principal (matriz intercelular) en forma de estructuras redondeadas ubicadas en pequeños grupos separados (condrias). Según el tipo y densidad de las fibras se distinguen tres grupos de cartílago: cartílago hialino, cartílago elástico y fibrocartílago. En un adulto, ninguno de los tipos de cartílago enumerados contiene vasos sanguíneos. El cartílago se nutre ya sea por difusión a través de la membrana que lo recubre (pericondrio) o directamente del líquido sinovial (cartílago hialino articular).
El desarrollo del cartílago comienza con la formación del pericondrio, pero el cartílago tiene una capacidad limitada para regenerarse. Sin pericondrio (cartílago hialino), no se produce la regeneración. El cartílago es altamente resistente a la presión, capaz de deformarse elásticamente y resiste la abrasión.
Cartílago hialino. El cartílago hialino preparado es de color blanco lechoso y translúcido. Por tanto, se parece al vidrio esmerilado. Este tipo de cartílago recubre la superficie interna de las articulaciones, forma los cartílagos costales y forma parcialmente el tabique nasal, la laringe, la tráquea y los bronquios grandes. Durante el período embrionario, la mayor parte del esqueleto se deposita en forma de cartílago. Con el crecimiento posterior del cuerpo, se forma cartílago hialino entre la epífisis (la parte en crecimiento del hueso) y el cuerpo del hueso, que es reemplazado por tejido óseo solo después de que se ha detenido el crecimiento. El cartílago hialino articular es el único tipo de cartílago que no contiene pericondrio. Por tanto, cuando se destruyen (como consecuencia de procesos inflamatorios o degenerativos en las articulaciones), no se produce una regeneración posterior.
Cartílago elástico. Junto con las estructuras presentes en el cartílago hialino, el cartílago elástico contiene una red ramificada de fibras elásticas que se localizan alrededor de los condrocitos y penetran en el pericondrio. Debido a la presencia de fibras elásticas, el cartílago presenta un color amarillento. En los seres humanos, el cartílago elástico se encuentra en el pabellón auricular, la epiglotis y el meato auditivo externo (canal auditivo).
Cartílago fibroso. A diferencia del cartílago hialino, el fibrocartílago contiene muchas más fibras de colágeno. El cartílago fibroso se localiza en zonas del esqueleto que suelen estar sometidas a carga debido a la acción de tendones y ligamentos. Se trata de discos intervertebrales (anillo fibroso), así como discos intraarticulares (discos y meniscos).
Tejidos conectivos- es un complejo de derivados mesenquimales, formado por diferencias celulares y una gran cantidad de sustancia intercelular (estructuras fibrosas y sustancia amorfa), implicados en el mantenimiento de la homeostasis del medio interno y que se diferencian de otros tejidos por una menor necesidad de procesos oxidativos aeróbicos. .
El tejido conectivo representa más del 50% del peso del cuerpo humano. Participa en la formación del estroma de los órganos, las capas entre otros tejidos, la dermis de la piel y el esqueleto.
El concepto de tejido conectivo (tejidos del medio interno, tejidos tróficos de soporte) combina tejidos que son diferentes en morfología y funciones, pero que tienen algunas propiedades comunes y se desarrollan a partir de una única fuente: el mesénquima.
Características estructurales y funcionales de los tejidos conectivos:
ubicación interna en el cuerpo;
predominio de la sustancia intercelular sobre las células;
variedad de formas celulares;
la fuente común de origen es el mesénquima.
Funciones de los tejidos conectivos:
mecánico;
apoyo y construcción de formas;
protector (mecánico, inmunológico inespecífico y específico);
reparador (plástico).
trófico (metabólico);
Morfogenético (formador de estructuras).
En realidad tejidos conectivos:
Tejidos conectivos fibrosos:
Tejido conectivo fibroso laxo informe
Desformado
Tejido conectivo fibroso denso:
Desformado
Decorado
Tejidos conectivos con propiedades especiales:
Tejido reticular
Tejido adiposo:
Mucoso
pigmentado
Tejido conectivo fibroso laxo informe
Peculiaridades:
Muchas células, poca sustancia intercelular (fibras y sustancia amorfa).
Localización:
forma el estroma de muchos órganos, la adventicia de los vasos, ubicada debajo de los epitelios; forma su propia lámina de membranas mucosas, submucosa, ubicada entre las células y fibras musculares
Funciones:
1. Función trófica: ubicada alrededor de los vasos, pvst regula el metabolismo entre la sangre y los tejidos del órgano.
2. La función protectora se debe a la presencia de macrófagos, células plasmáticas y leucocitos en el pvst. Los antígenos que atraviesan la barrera epitelial I del cuerpo se encuentran con la barrera II: células de defensa inespecífica (macrófagos, granulocitos neutrófilos) e inmunológicas (linfocitos, macrófagos, eosinófilos).
3. Función soporte-mecánica.
4. Función plástica: participa en la regeneración de órganos después de un daño.
Células (10 tipos)
1. Fibroblastos
Células fibroblásticas diferentes: células madre y semi-madre, fibroblastos poco especializados, fibroblastos diferenciados, fibrocitos, miofibroblastos, fibroclastos.
Células madre y semi-madre- Son pequeñas células de reserva cambial que rara vez se dividen.
Fibroblasto no especializado- células pequeñas, débilmente ramificadas con citoplasma basófilo (debido a la gran cantidad de ribosomas libres), orgánulos poco expresados; se divide activamente por mitosis, no participa de manera significativa en la síntesis de sustancia intercelular; como resultado de una mayor diferenciación, se convierte en fibroblastos diferenciados.
Fibroblastos diferenciados- las células funcionalmente más activas de esta serie: sintetizan proteínas de fibra (proelastina, procolágeno) y componentes orgánicos de la sustancia principal (glucosaminoglicanos, proteoglicanos). De acuerdo con su función, estas células tienen todas las características morfológicas de una célula que sintetiza proteínas: en el núcleo: nucléolos claramente definidos, a menudo varios; predomina la eucromatina; en el citoplasma: el aparato sintetizador de proteínas está bien expresado (EPS granular, complejo laminar, mitocondrias).
A nivel óptico de luz: células débilmente ramificadas con límites poco claros, con citoplasma basófilo; el núcleo es ligero, con nucléolos.
Hay 2 poblaciones de fibroblastos: De corta duración (varias semanas) Función:
protector. De corta duración (varias semanas) De larga duración (varios meses)
musculoesquelético. fibrocito
Las células de la serie fibroblástica son las células pvst más numerosas (hasta el 75% de todas las células) y producen la mayor parte de la sustancia intercelular.
El antagonista es fibroclasto- una célula con un gran contenido de lisosomas con un conjunto de enzimas hidrolíticas asegura la destrucción de la sustancia intercelular. Las células con alta actividad fagocítica e hidrolítica participan en la "resorción" de la sustancia intercelular durante el período de involución de los órganos (por ejemplo, el útero después del embarazo). Combinan las características estructurales de las células formadoras de fibrillas (retículo endoplásmico granular desarrollado, aparato de Golgi, mitocondrias relativamente grandes pero pocas), así como lisosomas con sus enzimas hidrolíticas características.
miofibroblasto- una célula que contiene proteínas actomiosina contráctiles en el citoplasma y, por tanto, capaz de contraerse. Células que son morfológicamente similares a los fibroblastos y combinan la capacidad de sintetizar no solo colágeno, sino también proteínas contráctiles en cantidades significativas.
Se ha establecido que los fibroblastos pueden transformarse en miofibroblastos, que son funcionalmente similares a las células del músculo liso, pero a diferencia de estas últimas tienen un retículo endoplasmático bien desarrollado. Estas células se observan en el tejido de granulación durante la cicatrización de heridas y en el útero durante el embarazo. Participan en la cicatrización de heridas, acercando los bordes de la herida durante la contracción.
2. Macrófagos
Funciones: Las siguientes células pvst en número son los macrófagos tisulares (sinónimo: histiocitos), que constituyen entre el 15 y el 20% de las células pvst. Se forman a partir de monocitos sanguíneos y pertenecen al sistema de macrófagos del cuerpo. Células grandes con núcleo polimórfico (redondo o en forma de frijol) y gran cantidad de citoplasma. De los orgánulos, los lisosomas y las mitocondrias están bien definidos. Contorno desigual de la citomembrana, capaz de movimiento activo.
función protectora mediante fagocitosis y digestión de partículas extrañas, microorganismos, productos de degradación de tejidos; participación en la cooperación celular en la inmunidad humoral; producción de la proteína antimicrobiana lisozima y de la proteína antiviral interferón, un factor que estimula la inmigración de granulocitos.
Constituyen el 10% de todas las células pvst. Suelen estar situados alrededor de los vasos sanguíneos. Célula redonda, ovalada, grande, a veces ramificada, con un diámetro de hasta 20 micrones; en el citoplasma hay muchos gránulos basófilos. Los gránulos contienen heparina e histamina, serotonina, quimasa y triptasa. Cuando se tiñen, los gránulos de mastocitos tienen la propiedad metacromasia- cambio de color del tinte. Los precursores de los basófilos tisulares se originan a partir de células madre hematopoyéticas de la médula ósea roja. Los procesos de división mitótica de los mastocitos se observan muy raramente.
Funciones: La heparina reduce la permeabilidad de las sustancias intercelulares y la coagulación sanguínea y tiene un efecto antiinflamatorio. La histamina actúa como su antagonista. La cantidad de basófilos tisulares varía según las condiciones fisiológicas del cuerpo: aumenta en el útero y las glándulas mamarias durante el embarazo, y en el estómago, los intestinos y el hígado en el momento álgido de la digestión. En general, los mastocitos regulan la homeostasis local.
4. Plasmocitos
Formado a partir de linfocitos B. En morfología son similares a los linfocitos, aunque tienen características propias. El núcleo es redondo y está ubicado excéntricamente; La heterocromatina se encuentra en forma de pirámides con un vértice afilado hacia el centro, delimitadas entre sí por franjas radiales de eucromatina; por lo tanto, el núcleo del plasmocito se arranca como una "rueda con radios". El citoplasma es basófilo, con un "patio" ligero cerca del núcleo. Bajo un microscopio electrónico, el aparato sintetizador de proteínas es claramente visible: EPS granular, complejo laminar (en el área del "patio" luminoso) y mitocondrias. El diámetro de la celda es de 7 a 10 micrones. De corta duración (varias semanas) Son células efectoras de la inmunidad humoral: producen anticuerpos específicos (gammaglobulinas).
5. Leucocitos
Los leucocitos liberados de los vasos siempre están presentes en el RVST.
6. Lipocitos (sinónimos: adipocito, célula grasa).
1). Lipocitos blancos- células redondas con una estrecha franja de citoplasma alrededor de una gran gota de grasa en el centro. Hay pocos orgánulos en el citoplasma. El pequeño núcleo está situado de forma excéntrica. Al preparar preparaciones histológicas de la manera habitual, se disuelve una gota de grasa en alcohol y se lava, de modo que la estrecha franja de citoplasma restante en forma de anillo con un núcleo ubicado excéntricamente se asemeja a un anillo.
De corta duración (varias semanas) Los lipocitos blancos almacenan grasa como reserva (material energético rico en calorías y agua).
2). Lipocitos marrones- células redondas con una ubicación central del núcleo. Las inclusiones grasas en el citoplasma se detectan en forma de numerosas gotitas pequeñas. El citoplasma contiene muchas mitocondrias con alta actividad de la enzima oxidativa citocromo oxidasa que contiene hierro (color marrón). De corta duración (varias semanas) Los lipocitos marrones no acumulan grasa, sino que, por el contrario, la "queman" en las mitocondrias, y el calor liberado en este caso se utiliza para calentar la sangre en los capilares, es decir, participación en la termorregulación.
7. Células adventicias
Se trata de células poco especializadas que acompañan a los vasos sanguíneos. Tienen forma aplanada o fusiforme con citoplasma débilmente basófilo, núcleo ovalado y una pequeña cantidad de orgánulos. Durante el proceso de diferenciación, estas células aparentemente pueden convertirse en fibroblastos, miofibroblastos y adipocitos.
8. Pericitos
Ubicado en el espesor de la membrana basal de los capilares; participar en la regulación de la luz de los hemocapilares, regulando así el suministro de sangre a los tejidos circundantes.
9. Células endoteliales vasculares
Se forman a partir de células mesenquimales poco diferenciadas y cubren todos los vasos sanguíneos y linfáticos desde el interior; producen muchas sustancias biológicamente activas.
10. Melanocitos (células pigmentarias, pigmentocitos)
Células procesadas con inclusiones de pigmento melanina en el citoplasma. Origen: de células que migran desde la cresta neural. De corta duración (varias semanas) Protección UV.
El tejido conectivo incluye tejido fibroso, tejido conectivo con propiedades especiales y tejido esquelético (cartílago y hueso). El tejido conectivo está formado por células y una gran cantidad de sustancia intercelular, que consta de fibras y sustancia fundamental.
El tejido conectivo fibroso incluye el denso laxo, el informe y el denso formado (tendones, membranas fibrosas, tejidos laminares y elásticos). El tejido conectivo con propiedades especiales está representado por tejido reticular, adiposo, mucoso y pigmentado.
El tejido conectivo realiza una función trófica asociada a la nutrición de las células y su participación en el metabolismo, protectora (fagocitosis, producción de cuerpos inmunes), mecánica (forma el estroma de los órganos, los conecta entre sí, forma fascia, etc.), plástica. (participa en procesos de regeneración, cicatrización de heridas) funciones. En algunas condiciones patológicas, el tejido conectivo puede participar en la hematopoyesis, ya que sus células pueden dar lugar a elementos sanguíneos.
Tejido conectivo fibroso laxo. Este tejido está formado por células y sustancia intercelular, en las que las fibras están ubicadas de forma suelta y tienen diferentes direcciones (Fig. 4). Acompaña a los vasos sanguíneos y nervios y forma parte de los órganos formando su estroma. La sustancia intercelular contiene colágeno (adhesivo), fibras elásticas y sustancia fundamental.
Arroz. 4. Tejido conectivo fibroso laxo. 1 - fibra de colágeno; 2 - fibras elásticas; 3 - macrofagocitos; 4 - fibroblastos; 5 - linfocitos
Las fibras de colágeno son hebras curvas, rectas u onduladas, de 1 a 12 micrones de espesor, que constan de hilos aún más delgados: fibrillas. Pueden hincharse y son muy duraderos. Las fibras elásticas son hilos ramificados de diferentes diámetros. Pueden detectarse mediante tinción especial de preparaciones histológicas. En el tejido conjuntivo fibroso laxo forman una red de bucles anchos. Además de estos dos tipos de fibras, el tejido conectivo laxo también contiene fibras reticulares o argirófilas, que reciben su nombre porque se tiñen fácilmente con sales de plata y forman una red. Forman parte del estroma de los ganglios linfáticos, bazo, médula ósea, etc.
La sustancia principal del tejido conectivo es una masa homogénea y es un coloide. Contiene mucopolisacáridos (ácido hialurónico, heparina, etc.), que determinan las características morfológicas y funcionales de la sustancia principal. Los elementos celulares del tejido conectivo están representados por células poco diferenciadas, fibroblastos, macrófagos (macrófagos), basófilos tisulares, plasmocitos, lipocitos y pigmentocitos. Además, las células sanguíneas (leucocitos) se encuentran en el tejido conectivo.
En un cuerpo adulto, las células cambian todo el tiempo. Las células moribundas son reemplazadas por otras nuevas debido a la reproducción de las de su propia especie. Además, el tejido conectivo contiene células que pueden transformarse en otras formas celulares. Estas células se denominan poco diferenciadas. Estos incluyen células ubicadas a lo largo de los capilares sanguíneos: adventiciales o perivasculares (pericitos). Lo mismo son las células reticulares y los linfocitos. Desempeñan un papel importante no solo en los procesos de restauración fisiológica del tejido, sino también en diversas condiciones patológicas (inflamación, trastornos hematopoyéticos, etc.).
Los fibroblastos son células planas con forma de huso, ampliamente representadas en el tejido conectivo. Son móviles y capaces de dividirse; pueden surgir de formas poco diferenciadas y transformarse en otras células.
Los fibroblastos participan en la formación de la sustancia fundamental y de las fibras de colágeno. En condiciones patológicas, participan en la cicatrización de heridas y en la formación de tejido cicatricial y cápsula de tejido conectivo alrededor de cuerpos extraños. Los fibroblastos que han completado su ciclo de desarrollo se denominan fibrocitos.
Los plasmocitos (células plasmáticas) se encuentran en el tejido conectivo laxo de la mucosa intestinal, el epiplón, varias glándulas, los ganglios linfáticos y la médula ósea. En algunas condiciones patológicas su número aumenta considerablemente. Tienen diferentes formas y tamaños y pueden surgir de linfocitos, células reticulares, macrófagos, etc. Las células plasmáticas intervienen en la formación de anticuerpos, así como en el metabolismo de las proteínas.
Los lipocitos (células grasas) tienen la capacidad de acumular grasa de reserva. Se encuentran en el tejido conectivo laxo solos o en grupos cerca de los vasos sanguíneos. Cuando los lipocitos se acumulan en gran cantidad desplazando a otras células, se habla de tejido adiposo.
Las células grasas son esféricas, normalmente cada célula contiene una gota de grasa neutra, ocupando toda la parte central de la célula. La cantidad de células grasas en el tejido conectivo varía mucho. Con mayor frecuencia se forman a partir de células adventicias que acompañan a los capilares sanguíneos. Los pigmentocitos (células pigmentarias) son células alargadas con procesos cortos y de forma irregular. Su citoplasma contiene granos del pigmento melanina. En el tejido conectivo laxo se encuentran en la piel alrededor. ano
, en la piel del escroto y pezones de las glándulas mamarias. Hay muchos de ellos en la coroides del ojo.

Tejido conectivo fibroso denso. Dependiendo de la disposición de las fibras, este tejido se divide en sin forma y decorado. Es imposible trazar un límite definido entre el tejido conectivo laxo y el denso y informe. En este último hay menos sustancia básica, fibras de colágeno y una red de fibras elásticas están estrechamente adyacentes entre sí, entrelazadas, asemejándose al fieltro. Contiene pocos elementos celulares. En el tejido conectivo fibroso denso formado, los haces de fibras de colágeno se ubican en una determinada dirección, correspondiente a las condiciones mecánicas en las que funciona el órgano (Fig. 5). Forma tendones musculares, ligamentos, membranas y tejido conectivo laminar que recubre algunos órganos (perineurio, cuerpos laminares, etc.). Algunos ligamentos (ligamentos amarillos de la columna vertebral, cuerdas vocales, etc.) y membranas de las paredes de órganos huecos y vasos sanguíneos están formados por tejido elástico que contiene una gran cantidad de fibras elásticas.
Tejido conectivo con propiedades especiales. El tejido reticular está formado por células reticulares y fibras reticulares. Las células reticulares tienen prolongaciones con las que se conectan entre sí formando una malla (retículo; de ahí el nombre del tejido). Las fibras reticulares se encuentran en todas direcciones. Las fibras reticulares se encuentran en todas direcciones. El tejido reticular forma la columna vertebral de la médula ósea, los ganglios linfáticos y el bazo, y también se encuentra en la mucosa intestinal, los riñones, etc. Las células reticulares pueden transformarse en células de otro tipo (hemocitoblastos, macrófagos, fibroblastos, etc.).
El sistema reticuloendotelial (sistema de macrófagos) es el conjunto de todas las células del organismo capaces de capturar partículas coloides y en suspensión de un medio líquido y depositarlas en el citoplasma. Estas células sirven para destruir agentes nocivos para el organismo, procedentes del exterior o que aparecen localmente, en el interior del organismo. Desempeñan un papel importante en la formación de inmunidad. Estas células incluyen macrófagos que fagocitan células reticulares de órganos hematopoyéticos, células estrelladas de capilares sanguíneos sinusoidales del hígado, etc. Por primera vez, I. I. Mechnikov unió estas células en un solo sistema.
El tejido adiposo es un lugar de acumulación de nutrientes de reserva, por lo que su cantidad cambia según la nutrición del organismo. En los seres humanos, el tejido adiposo forma una capa subcutánea, ubicada en el epiplón, el mesenterio del intestino, cerca de los riñones, etc. Por lo general, se divide en capas de tejido conectivo laxo en lóbulos. Las células grasas contienen gotitas de grasa y suelen tener forma esférica o poligonal. Entre ellos pasan fibras de colágeno y elásticas y se localizan fibroblastos, mastocitos y linfocitos. En el tejido adiposo tienen lugar procesos metabólicos activos, en particular la formación de grasas a partir de carbohidratos.
El tejido conectivo mucoso o gelatinoso se encuentra sólo en el embrión, en particular en el cordón umbilical humano. La sustancia intercelular de este tejido es homogénea y se parece a la gelatina.
El tejido pigmentado es un tejido que contiene muchas células pigmentarias: los melanocitos.
Tejido cartilaginoso. Este tejido está formado por células especiales: condrocitos, rodeados por una gran cantidad de sustancia intercelular. Dependiendo de la estructura de la sustancia intercelular, se distinguen el cartílago hialino, elástico y fibroso.
El cartílago hialino (Fig. 6) está formado por células cartilaginosas que se encuentran en cavidades especiales en la sustancia intercelular, generalmente en grupos. Células de diversas formas, con mayor frecuencia redondas u ovaladas. La sustancia intercelular es transparente y está formada por fibras de colágeno y sustancia fundamental. El cartílago en el cuerpo adulto forma la parte cartilaginosa de las costillas, cubre las superficies de los huesos articulados y forma el esqueleto. tracto respiratorio. Con la edad, hay una disminución en el número de células de cartílago y un cambio en la composición química de la sustancia intercelular, como resultado de lo cual se depositan sales de calcio y se produce la calcificación del cartílago.

El cartílago elástico en el ser humano forma el pabellón auricular, algunos cartílagos de la laringe, etc., tiene un color amarillento y es menos transparente que el hialino.
La sustancia intercelular contiene una gran cantidad de fibras elásticas.
En él nunca se produce el proceso de calcificación.
El cartílago fibroso forma los discos intervertebrales, la sínfisis púbica y recubre las superficies articulares de las articulaciones temporomandibular, esternoclavicular y algunas otras. Su sustancia intercelular contiene una gran cantidad de fibras de colágeno.

El pericondrio cubre la superficie del cartílago. Su capa interna contiene células especiales, condroblastos, a partir de los cuales se desarrollan las células del cartílago, los condrocitos, como resultado del crecimiento del cartílago.
El tejido óseo está en constante destrucción y creación de hueso.
Las propiedades fisiológicas del tejido óseo pueden cambiar con la edad, dependiendo de la nutrición, la actividad muscular y cuando se altera la actividad de las glándulas endocrinas y la inervación. Las fibras de colágeno del tejido óseo se llaman oseína (os - hueso); se revelan sobre preparaciones histológicas con un tratamiento especial. Las sustancias inorgánicas están representadas principalmente por sales de calcio, que forman compuestos complejos que dan fuerza a los huesos. La sustancia ósea orgánica, la oseína, hace que el hueso sea flexible y elástico. La combinación de estas propiedades crea la resistencia y ligereza necesarias para el tejido de soporte. En la sustancia intercelular del tejido óseo hay cavidades planas de forma ovalada, llamadas cavidades óseas. Están conectados por túbulos óseos. Hay tres tipos de células que se encuentran en el tejido óseo: osteoblastos, osteocitos y osteoclastos.
Los osteoblastos son células que forman tejido óseo. Se encuentran en lugares de destrucción y restauración del tejido óseo. Hay muchos de ellos en el hueso en desarrollo.
Los osteocitos se forman a partir de osteoblastos y tienen prolongaciones. Los cuerpos de los osteocitos se encuentran en las cavidades óseas y los procesos se extienden hasta los canalículos óseos. El sistema de túbulos óseos crea las condiciones para el intercambio de sustancias entre los osteocitos y el líquido tisular.
Los osteoclastos son grandes células multinucleadas con procesos. Participan en la destrucción de huesos y cartílagos calcificados con la formación de una bahía o laguna.
Hay dos tipos de tejido óseo: de fibra gruesa y laminar. También incluye la dentina dental.
En el tejido óseo grueso, las fibras de colágeno forman haces claramente visibles, entre los cuales se encuentran los osteocitos en las cavidades óseas. En los humanos, este tejido se encuentra solo durante el desarrollo de los huesos en el embrión, y en los adultos, en las suturas del cráneo y en los lugares de unión de los tendones a los huesos.
El tejido óseo laminar o fibroso fino contiene fibras de colágeno ubicadas en haces paralelos dentro de las placas o entre ellas. El tejido óseo laminar forma todos los huesos del esqueleto humano.
Hueso. El tejido óseo laminar forma la sustancia ósea compacta y esponjosa que forma el hueso. En la sustancia ósea compacta, las placas óseas están dispuestas en un orden determinado y confieren a la sustancia una mayor densidad (Fig. 8). En la sustancia esponjosa, las placas del interior del hueso forman barras transversales de diferentes formas, ubicadas según la función del hueso.

Arroz. 8. Estructura ósea. 1, 5 - canales centrales de osteona; 2 - placas de osteonas; 3 - placas de inserción; 4 - platos comunes
La sustancia compacta está formada principalmente por la parte media de los huesos tubulares largos (el cuerpo o diáfisis), y la sustancia esponjosa forma sus extremos, o epífisis, así como los huesos cortos; Los huesos planos contienen ambas sustancias.
En la sustancia ósea compacta, las placas óseas forman unos sistemas tubulares peculiares: las osteonas. Osteon es una unidad estructural del hueso.
Las placas óseas están dispuestas concéntricamente alrededor de los vasos sanguíneos;
Por lo general, hay de 5 a 20 con un espesor de 3 a 7 micrones. Este diseño confiere al hueso una resistencia especial. La cavidad en el centro del osteon, por donde pasa el vaso, se llama canal central del osteon (canal de Havers). Los canales se conectan entre sí y los vasos se conectan entre sí, con los vasos de la médula ósea ubicados dentro del hueso y con los vasos del periostio. Entre las osteonas, las placas óseas discurren en diferentes direcciones y se denominan intercalares o intermedias. Fuera y dentro del hueso, las placas están dispuestas de forma concéntrica. Los canales a través de los cuales pasan los vasos desde el periostio al hueso se denominan canales de nutrientes. El periostio está conectado al hueso mediante fibras de colágeno llamadas fibras perforantes o de Sharpey.
El exterior del hueso está cubierto de periostio (periostio). Consta de dos capas de tejido conectivo. La capa interna contiene muchas fibras elásticas y de colágeno, así como osteoclastos y osteoblastos. Durante el crecimiento, los osteoblastos del periostio también participan en la formación de hueso. La capa exterior está hecha de tejido conectivo más denso; a ella se unen ligamentos y tendones musculares. El periostio contiene una gran cantidad de vasos y nervios.
El tejido conectivo humano está formado por células inmóviles (fibrocitos, fibroblastos), que forman la sustancia fundamental y la sustancia intercelular fibrosa. Además, en el tejido conectivo (como en otros tejidos laxos) hay varias células libres (grasas, grasas, errantes, etc.). El tejido conectivo también incluye tejido óseo y cartilaginoso.
Funciones
Los tejidos conectivos, incluidos los de soporte (hueso, cartílago), dan forma, fuerza y estabilidad al cuerpo humano, y también protegen, cubren y conectan los órganos entre sí. La función principal de la sustancia intercelular es el apoyo y la sustancia principal asegura el intercambio de sustancias entre las células y la sangre.
tipos
- Embrionario (mesénquima): formado en el útero. Está formado por todo tipo de tejido conectivo, células musculares, células sanguíneas, etc.
- Reticular: consta de células reticulocitos que pueden acumular agua y actuar como fagocitos. Este tejido participa en la producción de anticuerpos, ya que se encuentra en todos los órganos del sistema linfático y forma la base de la médula ósea roja.
- Intersticial: es el tejido de soporte de los órganos, informe o difuso, suelto, que llena los espacios entre los órganos internos. Además de las células, el tejido intersticial contiene estructuras fibrosas.
- Elástico: contiene una gran cantidad de fibras de colágeno fuertes que se encuentran en los ligamentos, tendones y fascias que cubren los músculos.
- Graso: protege al cuerpo de la pérdida de calor; en los vertebrados se encuentra principalmente debajo de la piel, en el epiplón y entre los órganos internos, formando almohadillas suaves y elásticas. En los seres humanos, está representado por tejido adiposo blanco y marrón.
tejido cartilaginoso
Resistente a la presión, flexible y bastante suave. Está formado por células acuosas y sustancia intercelular. Según la naturaleza de la sustancia intercelular, el cartílago se divide en hialino, elástico y fibroso. El cartílago casi no tiene vasos sanguíneos ni nervios. El cartílago hialino es de color blanco azulado y contiene una gran cantidad de fibras de colágeno. Está cubierto de pericondrio, consta del esqueleto embrionario, cartílagos articulares y costales, la mayoría de los cartílagos de la laringe y la tráquea. El cartílago elástico de color amarillento contiene fibras elásticas; está formado por la parte cartilaginosa de la aurícula, la epiglotis, secciones de la pared del conducto auditivo externo, algunos cartílagos de la laringe y los cartílagos de los bronquios pequeños. El cartílago elástico carece de calcio. El cartílago fibroso contiene menos células que los dos primeros tipos de cartílago, pero contiene muchas más placas de colágeno. Se encuentra en los discos intervertebrales, meniscos y sínfisis del pubis.
Hueso
Consta de elementos celulares y sustancia intercelular mineralizada. Las sales minerales determinan la resistencia de los huesos. El contenido de calcio en los huesos disminuye con la falta de vitaminas, así como con el desequilibrio hormonal. Los huesos forman el esqueleto humano y, junto con las articulaciones, forman el sistema musculoesquelético.
Masaje
El masaje del tejido conectivo es una forma especial de masaje de zonas reflexogénicas. Las yemas de los dedos masajean lentamente la piel y el tejido conectivo subcutáneo, provocando una respuesta que mejora la circulación sanguínea en los tejidos y órganos afectados de una persona.
Capítulo 8. TEJIDO CONECTIVO
Capítulo 8. TEJIDO CONECTIVO
Los tejidos conectivos son grupo grande Los tejidos derivados del mesénquima, por regla general, son polidiferentes y con predominio de sustancia intercelular (estructuras fibrosas y un componente amorfo), participando en el mantenimiento de la constancia de la composición del medio interno del cuerpo.
El tejido conectivo representa más del 50% del peso del cuerpo humano. Participa en la formación del estroma de los órganos, las capas entre otros tejidos, la dermis de la piel y el esqueleto. La naturaleza multifuncional de los tejidos conectivos está determinada por la complejidad de su composición y organización.
Funciones de los tejidos conectivos. Los tejidos conectivos realizan diversas funciones: trófica, protectora, de soporte (biomecánica), plástica, morfogenética. La función trófica en el sentido amplio de la palabra está asociada con la regulación de la nutrición de diversas estructuras tisulares, con la participación en el metabolismo y el mantenimiento de la constancia de la composición del ambiente interno del cuerpo. En la implementación de esta función, el papel principal lo desempeña la sustancia principal a través de la cual se lleva a cabo el transporte de agua, sales y moléculas de nutrientes: el medio tampón integrador. La función protectora es proteger el cuerpo de influencias mecánicas no fisiológicas (daños) y neutralizar sustancias extrañas que provienen del exterior o se forman dentro del cuerpo. Esto lo proporciona la protección física (tejido óseo), así como la actividad fagocítica de los macrófagos y las células inmunocompetentes implicadas en las reacciones de la inmunidad celular y humoral. La función de soporte (biomecánica) la proporcionan principalmente el colágeno y las fibras elásticas, que forman la base fibrosa de todos los órganos, y la composición y propiedades fisicoquímicas de la sustancia intercelular de los tejidos esqueléticos (mineralización). Cuanto más densa es la sustancia intercelular, más importante es la función biomecánica de apoyo. La función plástica del tejido conectivo se expresa en la adaptación a las condiciones de vida cambiantes, la regeneración y la participación en la sustitución de defectos de los órganos cuando están dañados. Morfogenético
La función (formadora de estructuras) se manifiesta por la formación de complejos tisulares y la provisión de servicios generales. organización estructuralórganos (formación de cápsulas, tabiques intraorgánicos), la influencia reguladora de algunos de sus componentes sobre la proliferación y diferenciación de células de diversos tejidos.
Clasificación de los tejidos conectivos. Los tipos de tejido conectivo difieren en la composición y proporción de diferencias celulares, fibras, así como en las propiedades fisicoquímicas de la sustancia intercelular amorfa. Los tejidos conectivos se dividen en tejido conectivo propiamente dicho (tejido conectivo laxo y tejidos conectivos con propiedades especiales) y tejidos esqueléticos. Estos últimos, a su vez, se dividen en tres tipos de tejido cartilaginoso (hialino, elástico, fibroso), dos tipos de tejido óseo (reticulofibroso y laminar), así como cemento y dentina del diente (Esquema 8.1).
Histogénesis de los tejidos conectivos. La fuente de desarrollo del tejido conectivo es el mesénquima (del griego. mesos- promedio, enchima- masa de relleno). Este es uno de los rudimentos embrionarios (según algunas ideas, tejido embrionario), que es una parte suelta de la capa germinal media. Los elementos celulares del mesénquima se forman en el proceso de diferenciación del dermatoma, esclerotoma, capas visceral y parietal del esplancnotoma. Además, existe un ectomesénquima (neuromesénquima), que se desarrolla a partir de la cresta neural (placa ganglionar). A medida que el embrión se desarrolla, células de origen diferente al de otras células embrionarias migran al mesénquima.
Esquema 8.1. Clasificación de tejidos conectivos.
rudimentos, por ejemplo, células neuroblásticas diferentes, mioblastos del músculo esquelético, pigmentocitos, etc.
En consecuencia, a partir de una determinada etapa del desarrollo del embrión, el mesénquima es un mosaico de células que surgen de diferentes capas germinales y rudimentos de tejido embrionario. Sin embargo, morfológicamente, todas las células mesenquimales difieren poco entre sí, y sólo los métodos de investigación muy sensibles (generalmente inmunocitoquímicos, microscópicos electrónicos) revelan células de diferente naturaleza dentro del mesénquima. El mesénquima existe sólo en el período embrionario del desarrollo humano. Después del nacimiento, en el cuerpo humano sólo quedan células poco diferenciadas (pluripotentes) como parte del tejido conectivo fibroso laxo (células adventicias), que pueden diferenciarse de manera divergente en diferentes direcciones, pero dentro de un determinado sistema tisular.
Hay histogénesis embrionaria y postembrionaria de los tejidos conectivos. Durante el proceso de histogénesis embrionaria, el mesénquima adquiere las características de la estructura tisular antes de la formación de otros tejidos. Este proceso ocurre de manera diferente en diferentes órganos y sistemas y depende de su importancia fisiológica desigual en las diferentes etapas de la embriogénesis. En la diferenciación del mesénquima, se observa asincronía topográfica tanto en el embrión como en los órganos extraembrionarios, altas tasas de reproducción celular y formación de fibras. La histogénesis postembrionaria en condiciones fisiológicas normales ocurre más lentamente y tiene como objetivo mantener la homeostasis del tejido, la proliferación de células poco diferenciadas y el reemplazo de las células moribundas. Un papel importante en estos procesos lo desempeñan las interacciones intercelulares intratisulares, los factores inductores e inhibidores (integrinas, factores adhesivos intercelulares, cargas funcionales, hormonas, oxigenación, presencia de células poco diferenciadas).
Principios generales de organización del tejido conectivo. Los componentes principales del tejido conectivo son derivados celulares: estructuras fibrosas de tipo colágeno y elástico, una sustancia básica (amorfa) que desempeña el papel de un entorno metabólico amortiguador integrador y elementos celulares que crean y mantienen la proporción cuantitativa y cualitativa de la composición. de componentes no celulares.
La especificidad orgánica de los elementos celulares del tejido conectivo se expresa en el número, forma y proporción de los diferentes tipos de células, su metabolismo y funciones, óptimamente adaptadas a la función del órgano. La especificidad de los elementos celulares también se manifiesta por su interacción entre sí (ubicados individualmente, asociaciones celulares), las características de su estructura interna (composición de orgánulos, estructura nuclear, presencia de enzimas, etc.). La especificidad del tejido conectivo también se revela en la proporción de células de diversas diferencias y estructuras no celulares en diferentes partes del cuerpo. En el tejido conectivo fibroso laxo, las células de diversas diferencias y materia amorfa prevalecen sobre las fibras, y en el tejido conectivo denso, por el contrario, la mayor parte de su masa está formada por fibras.
8.1. TEJIDO CONECTIVO APROPIADO
8.1.1. Tejidos conectivos fibrosos
Tejido conectivo laxo
Tejido conectivo laxo (texto conectivo laxo) Se encuentra en todos los órganos, ya que acompaña a los vasos sanguíneos y linfáticos y forma el estroma de muchos órganos. A pesar de la presencia de características orgánicas, la estructura del tejido conectivo laxo en varios órganos es similar. Consiste en células de diversas determinaciones histogenéticas y sustancias intercelulares (fig. 8.1).

Arroz. 8.1. Tejido conectivo laxo:
A- preparación de película: 1 - fibroblasto; 2 - macrófagos; 3 - fibras de colágeno; 4 - fibras elásticas; 5 - linfocitos; b- fibroblasto; V- macrófago

Arroz. 8.2. Estructura ultramicroscópica de un fibroblasto en diferentes etapas de diferenciación (según N. A. Yurina y A. I. Radostina, con modificaciones): Fibroblastos: A- poco diferenciados; b- joven; V- maduro; GRAMO- fibrocito. 1 - núcleo; 2 - complejo de Golgi; 3 - mitocondrias; 4 - ribosomas y polirribosomas; 5 - retículo endoplásmico granular; 6 - fibrillas de colágeno
Células
Las principales células del tejido conectivo son fibroblastos(familia de células formadoras de fibrillas), macrófagos(familia), mastocitos, células adventicias, células plasmáticas, pericitos, células grasas, y leucocitos, migrando de la sangre; A veces pigmento
células.
fibroblastos(del lat. fibra- fibra, griego blastos- brote, rudimento) - células que sintetizan los componentes de la sustancia intercelular: proteínas (colágeno, elastina), proteoglicanos, glicoproteínas (ver Fig. 8.1; Fig. 8.2, 8.3).
Entre las células mesenquimales se encuentran las células madre que dan lugar a la diferenciación de los fibroblastos: células madre, células precursoras semi-madre, fibroblastos poco diferenciados (poco especializados), diferenciados (maduros, que funcionan activamente), fibrocitos (definitivos, las formas finales de células), así como miofibroblastos y fibroclastos. La función principal de los fibroblastos está asociada con la formación de la sustancia y fibras principales, la cicatrización de heridas, el desarrollo del tejido de granulación, la formación de una cápsula de tejido conectivo alrededor de un cuerpo extraño, etc. Morfológicamente, en este diferencial solo se pueden identificar células, comenzando con fibroblastos poco diferenciados. Estas últimas son células pequeñas procesadas con un núcleo redondo u ovalado y un pequeño nucléolo, citoplasma basófilo, rico en ARN. El tamaño de la celda no supera los 20-25 micrones. En el citoplasma de estas células se encuentra.
una gran cantidad de ribosomas libres (ver Fig. 8.2). El retículo endoplásmico y las mitocondrias están poco desarrollados. El complejo de Golgi está representado por grupos de tubos cortos y vesículas.
Fibroblastos maduros diferenciados Las preparaciones de mayor tamaño y extendidas sobre películas pueden alcanzar entre 40 y 50 micrones o más (ver Fig. 8.1). Estas son células que funcionan activamente. Sus núcleos son claros, ovalados, contienen 1-2 nucléolos grandes; el citoplasma es basófilo, con un retículo endoplásmico granular bien desarrollado, que en algunos lugares está en contacto con el plasmalema (ver Fig. 8.2, Fig. 8.3, A). El complejo de Golgi se distribuye en forma de cisternas y vesículas por toda la célula. Las mitocondrias y los lisosomas están moderadamente desarrollados.
En el citoplasma de los fibroblastos, especialmente en la parte periférica, hay microfilamentos de 5-6 nm de espesor que contienen proteínas como la actina y la miosina, lo que determina la capacidad de movimiento de estas células. El movimiento de los fibroblastos sólo es posible después de que se unen a las estructuras fibrilares de soporte (fibrina, fibras del tejido conectivo) con la ayuda de la fibronectina, una glicoproteína sintetizada por los fibroblastos y otras células que asegura la adhesión de células y estructuras no celulares. Durante el movimiento, el fibroblasto se aplana y su superficie puede aumentar 10 veces.
El plasmalema de los fibroblastos es una importante zona receptora que media los efectos de diversos factores reguladores. La activación de los fibroblastos suele ir acompañada de la acumulación de glucógeno y una mayor actividad de las enzimas hidrolíticas. La energía generada por el metabolismo del glucógeno se utiliza para sintetizar polipéptidos y otros componentes secretados por la célula.
La biosíntesis de proteínas de colágeno y elastina, proteoglicanos necesarios para la formación de la sustancia principal y las fibras, en los fibroblastos maduros se lleva a cabo de forma bastante intensiva, especialmente en condiciones de baja concentración de oxígeno. Los factores estimulantes de la biosíntesis de colágeno son los iones hierro, cobre, cromo, ácido ascórbico. Una de las enzimas hidrolíticas, la colagenasa, descompone el colágeno inmaduro dentro de las células, lo que aparentemente regula la intensidad de la secreción de colágeno a nivel celular.
Según su capacidad para sintetizar proteínas fibrilares, la familia de los fibroblastos incluye células reticulares del tejido conectivo reticular de los órganos hematopoyéticos, condroblastos y osteoblastos de tipos de tejido conectivo esquelético.
fibrocitos- formas definitivas de desarrollo de fibroblastos. Estas células tienen forma de huso con procesos en forma de alas. Contienen una pequeña cantidad de orgánulos, vacuolas, lípidos y glucógeno. La síntesis de colágeno y otras sustancias en los fibrocitos se reduce drásticamente.
Miofibroblastos- células morfológicamente similares a los fibroblastos, que combinan la capacidad de sintetizar no solo colágeno, sino también contractilidad

Arroz. 8.3. Fibroblastos, miofibroblastos y fibroclastos:
A- fibroblasto (preparación de A. I. Radostina): 1 - núcleo; 2 - retículo endoplásmico granular; 3 - ribosomas; 4 - complejo de Golgi; 5 - mitocondrias; 6 - plasma-molema; 7 - fibrillas de colágeno; b- miofibroblasto (preparación de A. B. Schechter): 1 - núcleo; 2 - retículo endoplásmico granular; 3 - ribosomas; 4 - complejo de Golgi; 5 - filamentos contráctiles; 6 - plasmalema
proteínas corporales en cantidades significativas (ver Fig. 8.3, b). Se ha establecido que los fibroblastos pueden convertirse en miofibroblastos, que son funcionalmente similares a las células del músculo liso, pero a diferencia de estas últimas tienen un retículo endoplasmático bien desarrollado. Estas células se observan en

Arroz. 8.3. Continuación:
V- fibroclasto (preparación de A. B. Shekhter): 1 - núcleo; 2 - retículo endoplásmico granular; 3 - ribosomas; 4 - lisosomas; 5 - fagolisosomas con fragmentos de fibrillas de colágeno. Microfotografías electrónicas. Ampliación 20.000
tejido de granulación en condiciones de regeneración del proceso de la herida y en el útero durante el desarrollo del embarazo.
fibroclastos- las células con alta actividad fagocítica e hidrolítica participan en la "resorción" de la sustancia intercelular (ver Fig. 8.3, c) durante el período de involución del órgano (por ejemplo, el útero después del final del embarazo). Combinan las características estructurales de las células formadoras de fibrillas (retículo endoplásmico granular desarrollado, complejo de Golgi, mitocondrias relativamente grandes pero pocas), así como lisosomas con sus enzimas hidrolíticas características. El complejo de enzimas que secretan fuera de la célula descompone la sustancia cementante de las fibras de colágeno, tras lo cual se produce la fagocitosis y la digestión intracelular del colágeno por las proteasas ácidas de los lisosomas.
Macrófagos(macrofagocitos) (del griego. macros- grande y largo, fagos- devorador) es una población celular heterogénea especializada del sistema de defensa del cuerpo. Hay dos grupos de macrófagos: libres y fijos. Los macrófagos libres incluyen macrófagos de tejido conectivo laxo o histiocitos; macrófagos de cavidades serosas; macrófagos de exudados inflamatorios; macrófagos alveolares de los pulmones. Los macrófagos pueden moverse por todo el cuerpo. El grupo de macrófagos fijos (residentes) está formado por macrófagos de médula ósea, tejido óseo y cartilaginoso (osteoclastos, condroclastos),

Arroz. 8.4. Macrófagos:
A- macrófagos del tejido conectivo subcutáneo de rata (micrografía teñida con hematoxilina férrica); b- macrófago (micrografía electrónica; preparación de A.I. Radostina, aumento 18.000): 1 - núcleo; 2 - lisosomas primarios; 3 - lisosomas secundarios; 4 - perfiles de túbulos del retículo endoplásmico; 5 - microcrecimientos de la capa periférica del citoplasma
Zenks, ganglios linfáticos (macrófagos dendríticos), macrófagos intraepidérmicos (células de Langerhans), macrófagos de las vellosidades placentarias (células de Hofbauer), macrófagos del sistema nervioso central (microglía).
El tamaño y la forma de los macrófagos varían según su estado funcional (fig. 8.4). Generalmente macrófagos, a excepción de algunos de ellos.
especies (células gigantes de cuerpos extraños, condro y osteoclastos), tienen un núcleo. Los núcleos de los macrófagos son pequeños, redondos, con forma de frijol o de forma irregular. Contienen grandes grupos de cromatina. El citoplasma es basófilo, rico en lisosomas, fagosomas (características distintivas) y vesículas pinocitóticas, contiene una cantidad moderada de mitocondrias, retículo endoplásmico granular, complejo de Golgi, inclusiones de glucógeno, lípidos y otros (ver Fig. 8.4, b). En el citoplasma de los macrófagos, se distingue una "periferia celular", que proporciona al macrófago la capacidad de moverse, atraer microprotuberancias del citoplasma y realizar endo y exocitosis. Directamente debajo del plasmalema hay una red de filamentos de actina con un diámetro de 5-6 nm. A través de esta red pasan microtúbulos con un diámetro de 20 nm y se unen al plasmalema. Los microtúbulos se dirigen radialmente desde el centro celular hacia la periferia celular y desempeñan un papel importante en los movimientos intracelulares de lisosomas, vesículas micropinocitóticas y otras estructuras. En la superficie de la membrana plasmática se encuentran receptores para células tumorales y glóbulos rojos, linfocitos T y B, antígenos, inmunoglobulinas y hormonas. La presencia de receptores de inmunoglobulinas determina la participación de los macrófagos en las reacciones inmunitarias (consulte el Capítulo 14).
Formas de manifestación de la función protectora de los macrófagos: 1) absorción y posterior descomposición o aislamiento de material extraño; 2) neutralizarlo al contacto directo; 3) transferencia de información sobre material extraño a células inmunocompetentes capaces de neutralizarlo; 4) proporcionar un efecto estimulante sobre otra población de células del sistema de defensa del cuerpo. Los macrófagos tienen orgánulos que sintetizan enzimas para la descomposición intracelular y extracelular de materiales extraños, antibacterianos y otras sustancias biológicamente activas (proteasas, hidrolasas ácidas, pirógenos, interferón, lisozima, etc.).
La cantidad de macrófagos y su actividad aumentan especialmente durante los procesos inflamatorios. Los macrófagos producen factores quimiotácticos para los leucocitos. La IL-1 secretada por los macrófagos es capaz de aumentar la adhesión de los leucocitos al endotelio, la secreción de enzimas lisosomales por los neutrófilos y su citotoxicidad y activa la síntesis de ADN en los linfocitos. Los macrófagos producen factores que activan la producción de inmunoglobulinas por parte de los linfocitos B y la diferenciación de los linfocitos T y B; Los factores antitumorales citolíticos, así como los factores de crecimiento que influyen en la reproducción y diferenciación de las células de su propia población, estimulan la función de los fibroblastos (consulte el Capítulo 14).
El contacto de los macrófagos con los antígenos aumenta drásticamente el consumo de glucosa, el metabolismo de los lípidos y la actividad fagocítica.
Los macrófagos se forman a partir de HSC, así como de promonocitos y monocitos (v. fig. 7.15). La renovación completa de los macrófagos del tejido conectivo fibroso laxo en animales de experimentación ocurre aproximadamente 10 veces más rápido que los fibroblastos.
Uno de los tipos de macrófagos son las células gigantes multinucleadas, que antes se llamaban "células gigantes extrañas".
“cuerpos extraños”, ya que pueden formarse, en particular, en presencia de un cuerpo extraño. Las células gigantes multinucleadas contienen de 10 a 20 núcleos o más y surgen por fusión de macrófagos mononucleares o por endomitosis sin citotomía. Según la microscopía electrónica, las células gigantes multinucleadas contienen un aparato secretor y sintético y una gran cantidad de lisosomas. El plasmalema forma numerosos pliegues.
Concepto del sistema de macrófagos. Este sistema incluye macrófagos (histiocitos) de tejido conectivo fibroso laxo, células estrelladas de vasos sinusoidales del hígado, macrófagos libres y fijos de órganos hematopoyéticos (médula ósea, bazo, ganglios linfáticos), macrófagos del pulmón, exudados inflamatorios (macrófagos peritoneales) , osteoclastos, células gigantes de cuerpos extraños y macrófagos gliales del tejido nervioso (microglia). Todos ellos son fagocitos activos. El material fagocitado sufre una escisión enzimática dentro de la célula (“fagocitosis completa”), por lo que se eliminan los agentes nocivos para el organismo que surgen localmente o penetran desde el exterior. Las células tienen receptores de inmunoglobulinas en su superficie y surgen de promonocitos de la médula ósea y monocitos de la sangre. A diferencia de estos fagocitos "profesionales", los fibroblastos, las células reticulares, las células endoteliales, los granulocitos neutrófilos, etc. tienen la capacidad de absorción facultativa. Pero estas células no forman parte del sistema macrófago.
I. I. Mechnikov fue el primero en llegar a la idea de que la fagocitosis, que surge en el proceso de evolución como una forma de digestión intracelular y se asigna a muchas células, es al mismo tiempo un importante mecanismo de protección. Justificó la viabilidad de combinarlos en un solo sistema y propuso llamarlo macrófago. El sistema de macrófagos es un poderoso aparato protector que participa en reacciones protectoras tanto generales como locales del cuerpo. En todo el organismo, el sistema de macrófagos está regulado tanto por mecanismos locales como por los sistemas nervioso y endocrino.
En los años 1930-1940. este sistema protector se llamó reticuloendotelial. Recientemente, se le ha denominado sistema fagocitario mononuclear, lo que, sin embargo, no lo caracteriza con precisión debido a que entre las células incluidas en este sistema también se encuentran las multinucleadas (osteoclastos).
Mastocitos(mastocitos, basófilos tisulares, mastocitos). Estos términos se refieren a células en cuyo citoplasma hay una granularidad específica, que recuerda a los gránulos de los granulocitos basófilos. Los mastocitos participan en la reducción de la coagulación sanguínea, aumentando la permeabilidad de la barrera hemato-tejida, en el proceso de inflamación, inmuno-
génesis, etc. En los seres humanos, los mastocitos se encuentran dondequiera que haya capas de tejido conectivo laxo. Especialmente hay muchos mastocitos en la pared del tracto digestivo, el útero, la glándula mamaria, el timo y las amígdalas. A menudo se ubican en grupos a lo largo de los vasos sanguíneos de la microvasculatura: capilares, arteriolas, vénulas y pequeños vasos linfáticos (fig. 8.5, A).

Arroz. 8.5. Mastocitos: A - en el tejido conectivo subcutáneo (micrografía): 1 - núcleo; 2 - gránulos metacromáticos en el citoplasma; b- diagrama de la estructura ultramicroscópica (según Yu. I. Afanasyev): 1 - núcleo; 2 - complejo de Golgi; 3 - lisosoma; 4 - mitocondrias; 5 - retículo endoplásmico; 6 - microvellosidades; 7 - gránulos heterogéneos; 8 - gránulos secretores en la sustancia intercelular.
La forma de los mastocitos es variada. Las células pueden tener forma irregular, ovaladas. A veces, estas células tienen procesos cortos y anchos, lo que se debe a su capacidad de realizar movimientos ameboides. En los seres humanos, el ancho de estas células oscila entre 4 y 14 micrones y su longitud hasta 22 micrones. Los núcleos celulares son relativamente pequeños, generalmente de forma redonda u ovalada, con cromatina densamente empaquetada. Hay numerosos gránulos en el citoplasma. El tamaño, la composición y la cantidad de gránulos varían. Su diámetro es de aproximadamente 0,3-1 micrones (Fig. 8.5, b). Una minoría de gránulos son lisosomas azurófilos que se tiñen ortocromáticamente. La mayoría de los gránulos se caracterizan por metacromasia y contienen heparina, ácidos condroitinsulfúricos tipo A y C, ácido hialurónico, histamina y serotonina. Los gránulos tienen una estructura reticulada, laminar, cristaloide y mixta.
Los orgánulos de los mastocitos (mitocondrias, complejo de Golgi, retículo endoplásmico) están poco desarrollados. En el citoplasma se encuentran diversas enzimas: proteasas, lipasas, fosfatasas ácidas y alcalinas, peroxidasa, citocromo oxidasa, ATPasa, etc. Sin embargo, la histidina descarboxilasa, que se utiliza para sintetizar histamina a partir de histidina, debe considerarse una enzima marcadora.
Los mastocitos son capaces de secretar y liberar sus gránulos. La degranulación de los mastocitos puede ocurrir en respuesta a cualquier cambio en las condiciones fisiológicas y la acción de patógenos. Sin embargo, la liberación de aminas biogénicas de los mastocitos también puede ocurrir mediante la secreción de componentes solubles (secreción de histamina). La histamina provoca inmediatamente la dilatación de los capilares sanguíneos y aumenta su permeabilidad, lo que se manifiesta por edema local. También tiene propiedades hipotensoras y es un importante mediador de la inflamación.
La histamina actúa como antagonista de la heparina, lo que reduce la permeabilidad de la sustancia intercelular y la coagulación sanguínea y tiene un efecto antiinflamatorio.
La cantidad de mastocitos cambia según las condiciones fisiológicas del cuerpo: aumenta en el útero y las glándulas mamarias durante el embarazo, y en el estómago, los intestinos y el hígado en el momento álgido de la digestión.
Los precursores de los mastocitos se originan a partir de células madre hematopoyéticas de la médula ósea roja. Los procesos de división mitótica de los mastocitos se observan muy raramente.
Células plasmáticas (plasmocitos). Estas células aseguran la producción de anticuerpos: gammaglobulinas (proteínas) cuando aparece un antígeno en el cuerpo. Se forman en los órganos linfoides a partir de linfocitos B (ver Capítulo 14), que generalmente se encuentran en el tejido conectivo laxo de la lámina propia de las membranas mucosas de los órganos huecos, el epiplón, el tejido conectivo intersticial de varias glándulas (mamarias, salivales, etc.). ), ganglios linfáticos, bazo, médula ósea, etc.
El tamaño de las células plasmáticas oscila entre 7 y 10 micrones. La forma de las células es redonda u ovalada. Los granos son relativamente pequeños, de forma redonda u ovalada y están ubicados de forma excéntrica. El citoplasma es marcadamente basófilo y contiene endoplasma granular bien desarrollado ubicado concéntricamente.

Arroz. 8.6. Adipocitos:
1 - capilar; 2 - inclusiones de lípidos; 3 - núcleo; 4 - citoplasma (micrografía). Tinción - hematoxilina de hierro
Red tic en la que se sintetizan proteínas (anticuerpos). La basofilia está ausente sólo en una pequeña zona clara del citoplasma cerca del núcleo, formando la llamada esfera o patio. Aquí se encuentran los centríolos y el complejo de Golgi. A veces se encuentran acumulaciones de inmunoglobulinas en forma de oxifílicas. Corpúsculo de Roussel.
Las células plasmáticas se caracterizan por una alta tasa de síntesis y secreción de anticuerpos, lo que las distingue de sus predecesoras. Un aparato secretor bien desarrollado permite la síntesis y secreción de varios miles de moléculas de inmunoglobulina por segundo. El número de células plasmáticas aumenta en diversas enfermedades infecciosas, alérgicas e inflamatorias.
Adipocitos (células grasas). Se denomina así a las células que tienen la capacidad de acumular grandes cantidades de grasa de reserva, que interviene en el trofismo, la producción de energía y el metabolismo del agua. Los adipocitos se encuentran en grupos, con menos frecuencia solos y, por regla general, cerca de los vasos sanguíneos. Al acumularse en grandes cantidades, estas células forman tejido adiposo (blanco o marrón) (ver más abajo).
La forma de las células grasas individuales es esférica. Una célula grasa madura suele contener una gran gota de grasa neutra (triglicéridos), que ocupa toda la parte central de la célula y está rodeada por un fino borde citoplasmático, en cuya parte engrosada se encuentra el núcleo (fig. 8.6). Además, el citoplasma de los adipocitos contiene una pequeña cantidad de otros lípidos: colesterol, fosfolípidos, ácidos grasos libres, etc. Los lípidos se tiñen bien con Sudán III en naranja o con ácido ósmico en negro. En el citoplasma adyacente al núcleo y, a veces, en la parte opuesta más delgada del mismo.

Arroz. 8.7. Pigmentocitos en la piel (micrografía). La preparación no tiene color.
Se revelan mitocondrias filamentosas y en forma de bastón con crestas densamente ubicadas. En la periferia celular se encuentran numerosas vesículas pinocitóticas.
Los adipocitos del tejido adiposo marrón se distinguen por pequeñas inclusiones lipídicas y mitocondrias ubicadas alrededor del núcleo. Los adipocitos tienen una gran capacidad metabólica. Tanto el número de inclusiones grasas en los adipocitos como el número de células grasas en el tejido conectivo laxo están sujetos a fluctuaciones significativas.
Las funciones de los adipocitos son tróficas, asociadas al suministro de energía y reserva de agua en el organismo, así como a la participación en el proceso de termorregulación.
Se pueden desarrollar nuevas células grasas en el tejido conectivo del cuerpo adulto con una mayor nutrición de las células adventicias (cambiales) adyacentes a los capilares sanguíneos. En este caso, primero aparecen pequeñas gotas de grasa en el citoplasma de las células que, al aumentar de tamaño, se fusionan gradualmente en gotas más grandes. A medida que aumenta la caída de grasa, el retículo endoplásmico y el complejo de Golgi se reducen y el núcleo se comprime, aplana y empuja hacia la periferia de la célula.
Los grupos de células grasas que se encuentran en muchos órganos se denominan tejido adiposo.
Células adventicias. Estas son células poco diferenciadas (cambiales) que acompañan a los vasos sanguíneos pequeños. Tienen forma aplanada o fusiforme con citoplasma débilmente basófilo, núcleo ovalado y una pequeña cantidad de orgánulos. Estas células, mediante diferenciación divergente, dan lugar a diversas diferenciaciones celulares (fibroblásticas, miofibroblásticas, adipocíticas, etc.).
Pericitos- células que rodean los capilares sanguíneos y forman sus paredes (ver Capítulo 13).
pigmentocitos(células pigmentarias, melanocitos). Estas células contienen el pigmento melanina en su citoplasma. Hay muchos de ellos en las marcas de nacimiento, así como en el tejido conectivo de las personas de raza negra y amarilla. Los pigmentocitos tienen prolongaciones cortas de forma irregular (fig. 8.7), una gran cantidad de melanosomas (gránulos de melanina) que miden entre 15 y 25 nm y ribosomas. Algunos melanosomas de los melanocitos migran a los queratinocitos de las capas espinosa y basal de la epidermis.
El citoplasma de los melanocitos también contiene aminas biológicamente activas que pueden participar, junto con los mastocitos, en la regulación del tono de las paredes vasculares (ver Capítulo 14).
Los melanocitos sólo pertenecen formalmente al tejido conectivo, ya que se encuentran en él. En cuanto a su origen, se ha comprobado la formación de estas células a partir de células madre de las crestas neurales, y no a partir del mesenquima. Las células del tejido conectivo están funcionalmente conectadas en un solo sistema debido a numerosos factores de interacción, especialmente en los procesos de inflamación y regeneración postraumática, en violación del régimen agua-sal del cuerpo, etc.
Sustancia intercelular
Sustancia intercelular, o matriz (sustancia intercelular), El tejido conectivo está formado por fibras de colágeno y elásticas, así como por una sustancia fundamental (amorfa). La sustancia intercelular tanto en embriones como en adultos se forma, por un lado, mediante la secreción realizada por las células del tejido conectivo y, por otro, a partir del plasma sanguíneo que ingresa a los espacios intercelulares.
En los embriones humanos, la formación de sustancia intercelular se produce a partir del primer y segundo mes de desarrollo intrauterino. Durante la vida, la sustancia intercelular se renueva constantemente.
Las estructuras de colágeno que forman los tejidos conectivos de los organismos humanos y animales son sus componentes más representativos, formando una compleja jerarquía organizativa. La base de todo el grupo de estructuras de colágeno es la proteína fibrosa. colágeno, que determina las propiedades de las estructuras de colágeno.
Fibras de colágeno (fibras colagenae) como parte de diferentes tipos El tejido conectivo determina su fuerza. En el tejido conectivo laxo y informe, se ubican en diferentes direcciones en forma de hebras onduladas, curvadas, retorcidas en espiral, redondas o aplanadas con un espesor de 1 a 3 micrones o más. Su longitud es diferente. La estructura interna de la fibra de colágeno está determinada por la proteína fibrilar, el colágeno, que se sintetiza en los ribosomas del retículo endoplásmico granular de los fibroblastos.
Hay alrededor de 20 tipos de colágeno, que se diferencian en la organización molecular y la afiliación de órganos y tejidos.

Arroz. 8.8. Biosíntesis de colágeno y fibrilogénesis.
En el cuerpo humano los más comunes son los siguientes: el colágeno tipo I se encuentra principalmente en el tejido conectivo de la piel, tendones, huesos, córnea, esclerótica y paredes arteriales; El colágeno tipo II forma parte del cartílago hialino y fibroso, del cuerpo vítreo y de la córnea; El colágeno tipo III se encuentra en la dermis de la piel fetal, en las paredes de los grandes vasos sanguíneos y en las fibras reticulares de los órganos hematopoyéticos; Tipo IV: en las membranas basales, cápsula del cristalino; El colágeno tipo V está presente en el corion, amnios, endomisio, perimisio, piel, alrededor de las células (fibroblastos, endoteliales, músculo liso) que sintetizan colágeno.
Las moléculas de colágeno miden aproximadamente 280 nm de largo y 1,4 nm de ancho. Están formados por tripletes, tres cadenas polipeptídicas (cadenas α) del precursor del colágeno, el procolágeno, retorcidas en una sola espiral en la célula. Este es el primero molecular, Nivel de organización de las fibras de colágeno. El procolágeno se secreta en la sustancia intercelular (fig. 8.8).
Segundo, supramolecular, El nivel (organización extracelular de la fibra de colágeno) son moléculas de tropocolágeno agregadas en longitud y entrecruzadas mediante enlaces de hidrógeno, formados por la escisión de péptidos de procolágeno terminales. Primero, se forman protofibrillas, y 5-6 protofibrillas, conectadas entre sí por enlaces laterales, forman microfibrillas con un espesor de aproximadamente 5 nm.
Con la participación de glucosaminoglicanos, también secretados por fibroblastos, se forma el tercero, fibrilar, Nivel de organización de las fibras de colágeno. Las fibrillas de colágeno tienen estrías cruzadas.

Arroz. 8.9. Fibrilla de colágeno (preparación de N. P. Omelyanenko). P - punto. Micrografía electrónica. Aumentar 200.000
ny estructuras con un espesor promedio de 20-100 nm. El período de repetición de áreas oscuras y claras es de 64 a 67 nm (Fig. 8.9, 8.10). Se cree que cada molécula de colágeno en las filas paralelas está desplazada en un cuarto de la longitud con respecto a la cadena vecina, lo que provoca franjas oscuras y claras alternas. En las bandas oscuras bajo el microscopio electrónico se ven líneas transversales finas secundarias, causadas por la disposición de los aminoácidos polares en las moléculas de colágeno.
Cuatro, fibra, nivel de organización. La fibra de colágeno, formada por agregación de fibrillas, tiene un espesor de 1 a 10 μm (dependiendo de la topografía). Incluye una cantidad variable de fibrillas, desde una hasta varias docenas. Las fibras se pueden plegar en haces de hasta 150 micras de espesor.
Las fibras de colágeno se caracterizan por un bajo alargamiento y una alta resistencia a la tracción. En agua, el grosor del tendón aumenta en un 50% como resultado de la hinchazón, y en ácidos y álcalis diluidos, 10 veces, pero la fibra se acorta en un 30%. La capacidad de hincharse es más pronunciada en las fibras jóvenes. Cuando se tratan térmicamente en agua, las fibras de colágeno forman una sustancia adhesiva (griego: cola- pegamento), que dio nombre a estas fibras.
Los tipos de fibras de colágeno son reticular Y fibras precolágenas. Estos últimos representan la forma inicial de formación de fibras de colágeno en la embriogénesis y durante la regeneración. Contienen colágeno tipo III y una mayor cantidad de carbohidratos, que son sintetizados por las células reticulares de los órganos hematopoyéticos. Forman una red tridimensional: el retículo, que determina su nombre.
Fibras elásticas. Presencia de fibras elásticas. (fibra elástica) en el tejido conectivo determina su elasticidad y extensibilidad. Las fibras elásticas tienen una resistencia inferior a las fibras de colágeno. La forma de la sección transversal de las fibras es redonda y aplanada. En el tejido conectivo laxo se anastomosan ampliamente entre sí. El grosor de las fibras elásticas suele ser menor que el del colágeno (0,2-1 micrones), pero puede alcanzar varios micrómetros (por ejemplo, en el ligamento nucal). La composición de las fibras elásticas se divide en microfibrilar Y amorfo Componentes.

Arroz. 8.10. Formación de sustancia intercelular (según R. Krstic, con modificaciones):
1 - fibroblasto; 2 - cadenas polipeptídicas; 3 - moléculas de tropocolágeno; 4 - glucosaminoglicanos; 5 - polimerización de moléculas de tropocolágeno; 6 - protofibrilla; 7 - haz de protofibrillas (fibrilla de colágeno); 8 - molécula de elastina; 9 - fibrilla elástica
La base de las fibras elásticas es la glicoproteína globular. elastina, sintetizado por fibroblastos y células de músculo liso (primero, nivel de organización molecular). La elastina se caracteriza por un alto contenido de prolina y glicina y la presencia de dos derivados de aminoácidos: desmosina e isodesmosina, que participan en la estabilización de la estructura molecular de la elastina y le confieren capacidad de estiramiento y elasticidad. Las moléculas de elastina, que tienen glóbulos con un diámetro de 2,8 nm, están conectadas fuera de la célula en cadenas: protofibrillas de elastina con un espesor de 3-3,5 nm (segundo nivel de organización, supramolecular). Las protofibrillas de elastina en combinación con glicoproteína (fibrilina) forman microfibrillas (fig. 8.11) con un espesor de 8 a 19 nm (tercero, fibrilar, nivel de organización). El cuarto nivel de organización es

Arroz. 8.11. Estructura ultramicroscópica de fibra elástica: A- diagrama: 1 - parte central homogénea; 2 - microfibrillas (según Yu. I. Afanasyev); b- micrografía electrónica, aumento 45.000 (preparación de V. P. Slyusarchuk): 1 - parte central homogénea; 2 - microfibrillas en la periferia de la fibra; 3 - complejo de fibroblastos de Golgi; 4 - retículo endoplásmico; 5 - centríolo
fibra. Las fibras elásticas más maduras contienen aproximadamente el 90% del componente proteico elástico amorfo (elastina) en el centro y microfibrillas a lo largo de la periferia. En las fibras elásticas, a diferencia de las fibras de colágeno, no existen estructuras con estrías transversales a lo largo de su longitud.
Además de las fibras elásticas maduras, existen elaunina Y oxitalano fibras. En las fibras de elaunina, la proporción de microfibrillas con respecto al componente amorfo es aproximadamente igual, mientras que las fibras de oxitalano constan únicamente de microfibrillas.
Componente amorfo de la sustancia intercelular. Las células y fibras del tejido conectivo están encerradas en un componente amorfo o sustancia fundamental. (sustancia fundamental). Esta sustancia similar a un gel es un entorno multicomponente metabólico, amortiguador integrador que rodea las estructuras celulares y fibrosas del tejido conectivo, los nervios y los vasos sanguíneos. Los componentes de la sustancia principal incluyen proteínas del plasma sanguíneo, agua, iones inorgánicos, productos del metabolismo celular, precursores solubles de colágeno y elastina, proteoglicanos, glicoproteínas y complejos formados por ellos. Todas estas sustancias están en constante movimiento y renovación (fig. 8.12).
Los proteoglicanos (PG) son compuestos de proteínas y carbohidratos que contienen entre un 90 y un 95 % de carbohidratos.
Los glucosaminoglicanos (GAG) son compuestos polisacáridos que normalmente contienen ácido hexurónico con aminoazúcares (N-acetilglicosamina, ácido N-acetilgaláctico)
proteoglicano

Arroz. 8.12. Organización molecular de la sustancia amorfa del tejido conectivo.
tosamina). Las moléculas de GAG contienen muchos grupos hidroxilo, carboxilo y sulfato que tienen carga negativa, unen fácilmente moléculas de agua e iones, en particular Na +, y por lo tanto determinan las propiedades hidrófilas del tejido. Los GAG son permeables al oxígeno y al CO 2, pero protegen los órganos de la penetración de cuerpos extraños y proteínas. Los GAG participan en la formación de estructuras fibrosas del tejido conectivo y sus propiedades mecánicas, procesos reparadores del tejido conectivo y en la regulación del crecimiento y diferenciación celular. Entre estos compuestos, el más común en tipos de tejido conectivo es el ácido hialurónico, así como los GAG sulfatados: sulfatos de condroitina (en cartílago, piel, córnea), sulfato de dermatán (en piel, tendones, en la pared de los vasos sanguíneos, etc.). ), sulfato de queratán, sulfato de heparina (en la composición de muchas membranas basales).
La heparina es un GAG que consta de ácido glucurónico y glucosamina. En humanos y animales, es producido por mastocitos y es un factor anticoagulante natural en la sangre.
Las glicoproteínas (GP) son una clase de compuestos proteicos con oligosacáridos (hexosas, hexosas, fucosas, ácidos siálicos), que forman parte tanto de fibras como de sustancias amorfas. Estos incluyen GP solubles asociados con proteoglicanos; GP de tejidos calcificados; Médicos de cabecera relacionados con el colágeno. Los médicos de cabecera desempeñan un papel importante en la formación de la estructura de la sustancia intercelular del tejido conectivo y determinan sus características funcionales (fibronectina, condronectina, fibrilina, laminina, etc.).
La fibronectina es el principal GP de superficie de los fibroblastos. En el espacio intercelular se asocia principalmente con colágeno intersticial. Se cree que la fibronectina determina la pegajosidad, la motilidad, el crecimiento y la especialización de las células, etc.
La fibrilina forma microfibrillas y fortalece la conexión entre los componentes extracelulares.
La laminina es un componente de la membrana basal, que consta de tres cadenas polipeptídicas interconectadas por compuestos disulfuro, así como con colágeno tipo V y receptores de superficie celular.
Tejidos conectivos densos
Tejidos conectivos densos (texto conectivo compacto) caracterizado por una cantidad relativamente grande de fibras densamente ubicadas y una pequeña cantidad de elementos celulares y la principal sustancia amorfa entre ellos. Dependiendo de la naturaleza de la ubicación de las estructuras fibrosas, se distinguen. desorientado(desformado), orientado(formateado) y mezclado tipos de tejido conectivo denso.
Desorientado un tipo de tejido caracterizado por una disposición desordenada de fibras (por ejemplo, la dermis de la piel). EN orientado Dependiendo del tipo de tejido, la disposición de las fibras está estrictamente ordenada y corresponde en cada caso a las condiciones en las que funciona el órgano (tendones, ligamentos, membranas fibrosas). Mezclado El tipo, por regla general, tiene una estructura en capas (córnea, esclerótica) con direcciones alternas de elementos fibrosos de tejido conectivo denso.
Tendón(tendo). Consiste en haces paralelos de fibras de colágeno, gruesos y muy compactos, dispuestos en paralelo. Entre estos haces se encuentran fibrocitos llamados células del tendón (tendinocitos), y una pequeña cantidad de fibroblastos y material amorfo triturado. Los finos procesos laminares de fibrocitos ingresan a los espacios entre los haces de fibras y están en estrecho contacto con ellos. Cada haz de fibras de colágeno, separado de la capa adyacente de fibrocitos, se llama una viga de primer orden. Varios haces de primer orden, rodeados por finas capas de tejido conectivo laxo, forman los haces de segundo orden. Capas de tejido conectivo laxo que se separan vigas de segundo orden, son llamados endotenio. A partir de vigas de segundo orden se componen. vigas de tercer orden, separados por capas más gruesas de tejido conectivo laxo - peritenonio. A veces, el haz de tercer orden es el propio tendón. Los tendones grandes también pueden tener haces de cuarto orden.
El peritenonio y el endotenonio contienen vasos sanguíneos que irrigan el tendón, nervios y terminaciones nerviosas propioceptivas que envían señales al sistema nervioso central sobre el estado de tensión en el tejido del tendón.
El ligamento nucal también pertenece al tipo formado de tejido conectivo denso, solo que está formado a partir de fibras elásticas gruesas.
Algunos tendones en sus inserciones a los huesos están encerrados en vainas hechas de dos vainas de tejido conectivo fibroso. (vagina), entre los cuales hay un líquido (lubricante) rico en ácido hialurónico.
Membranas fibrosas. Estas estructuras, construidas a partir de tejido conectivo denso, incluyen fascia, aponeurosis, centros tendinosos del diafragma, cápsulas de algunos órganos, duramadre, esclerótica, pericondrio, periostio, así como la túnica albugínea del ovario y testículo, etc. Las membranas fibrosas de este tipo de tejido son difíciles de estirar debido a que los haces de fibras de colágeno y fibroblastos y fibrocitos que se encuentran entre ellas están dispuestos en un cierto orden en varias capas una encima de la otra. En cada capa, los haces ondulados de fibras de colágeno discurren paralelos entre sí en una dirección, que no coincide con la dirección en las capas adyacentes (tejido conectivo organizado en paralelo en diferentes direcciones). Haces individuales de fibras pasan de una capa a otra, conectándolas entre sí. Además de haces de fibras de colágeno, las membranas fibrosas contienen fibras elásticas. Las estructuras fibrosas como el periostio, la esclerótica, la túnica albugínea testicular, las cápsulas articulares y otras se caracterizan por una disposición menos regular de los haces de fibras de colágeno y una gran cantidad de fibras elásticas en comparación con las aponeurosis.
8.1.2. Tejidos conectivos con propiedades especiales.
Tales tejidos incluyen reticular, graso Y membrana mucosa Se caracterizan por un predominio de células homogéneas, a las que se suele asociar el propio nombre de este tipo de tejido conectivo.
Tejido reticular
Tejido reticular (texto reticular) es un tipo de tejido conectivo, tiene una estructura en forma de red y consta de procesos células reticulares Y fibras reticulares (argirófilas)(Figura 8.13). La mayoría de las células reticulares están asociadas con fibras reticulares y están conectadas entre sí mediante procesos, formando una red tridimensional. El tejido reticular forma el estroma de los órganos hematopoyéticos y el microambiente para las células sanguíneas que se desarrollan en ellos.
Las fibras reticulares (diámetro de 0,5 a 2 micrones) son un producto de la síntesis de células reticulares. Se detectan al impregnarse con sales de plata, por eso también se les llama argirofílicos (del griego. argentino- plata). Estas fibras son resistentes a los ácidos y álcalis débiles y no son digeridas por la tripsina.
En el grupo de las fibras argirófilas se distingue entre fibras reticulares y precolágenas. Las propias fibras reticulares son formaciones definitivas y finales que contienen colágeno tipo III. Las fibras reticulares, en comparación con las fibras de colágeno, contienen altas concentraciones de azufre, lípidos y carbohidratos. En un microscopio electrónico, las fibrillas de fibras reticulares no siempre tienen estrías claramente definidas con un período de 64-67 nm. En términos de extensibilidad, estas fibras ocupan
Tienen una posición intermedia entre el colágeno y el elástico. Las fibras de precolágeno representan la forma inicial de formación de fibras de colágeno en la embriogénesis y durante la regeneración.

Arroz. 8.13. Tejido reticular: A- micrografía de células reticulares: 1 - núcleo de una célula reticular; 2 - procesos del citoplasma; b- microfotografía de las fibras reticulares del ganglio linfático: 1 - fibras reticulares. Impregnación con nitrato de plata.
Tejido adiposo
Tejido adiposo (texto adiposo)- son acumulaciones de células grasas que se encuentran en muchos órganos (ver Adipocitos). Hay dos tipos de tejido adiposo: blanco Y marrón. Estos términos son condicionales y reflejan las características de la coloración celular. El tejido adiposo blanco está muy extendido en el cuerpo humano y el tejido adiposo marrón se encuentra principalmente en los recién nacidos y en algunos animales (roedores e hibernadores) durante toda la vida.
Tejido adiposo blanco en el ser humano se localiza debajo de la piel, especialmente en la parte inferior de la pared abdominal, en las nalgas y muslos, donde forma la capa de grasa subcutánea, en el epiplón, mesenterio y región retroperitoneal. El tejido adiposo está dividido más o menos claramente por capas de tejido conectivo fibroso laxo en lóbulos de diversos tamaños y formas. Las células grasas del interior de los lóbulos están bastante cerca unas de otras (fig. 8.14). En los estrechos espacios entre ellos se encuentran fibroblastos, elementos linfoides y mastocitos. Las finas fibras de colágeno están orientadas entre las células grasas en todas direcciones. Los capilares sanguíneos y linfáticos, ubicados en capas de tejido conectivo fibroso laxo, rodean estrechamente grupos de células grasas o lóbulos de tejido adiposo con sus asas. En el tejido adiposo se producen procesos activos de metabolismo de ácidos grasos, carbohidratos y formación de grasas a partir de carbohidratos.
Durante la descomposición de las grasas, se libera una gran cantidad de agua y energía, por lo que el tejido adiposo desempeña no solo el papel de depósito de sustratos para la síntesis de compuestos de alta energía, sino también indirectamente el papel de depósito de agua. .
Durante el ayuno, el tejido adiposo subcutáneo y perinéfrico, el tejido adiposo omental y mesentérico pierden rápidamente reservas de grasa. gotitas de lipi
Las grasas dentro de las células se trituran y las células grasas adquieren una forma estrellada o fusiforme. En la zona orbitaria de los ojos y en la piel de las palmas y las plantas, el tejido adiposo pierde sólo una pequeña cantidad de lípidos incluso durante un ayuno prolongado. En este caso, el tejido adiposo desempeña un papel predominantemente mecánico más que metabólico. En estos lugares se divide en pequeños lóbulos rodeados de fibras de tejido conectivo.
Tejido adiposo marrón Se encuentra en los recién nacidos y en algunos animales en el cuello, cerca de los omóplatos, detrás del esternón, a lo largo de la columna, debajo de la piel y entre los músculos. Consiste en células grasas densamente entrelazadas con hemocapilares. Estas células participan en los procesos de producción de calor. Los adipocitos del tejido adiposo marrón tienen muchas pequeñas inclusiones grasas en el citoplasma (fig. 8.15). En comparación con las células del tejido adiposo blanco, tienen muchas más mitocondrias. El color marrón de las células grasas lo dan los pigmentos que contienen hierro: los citocromos mitocondriales.
La capacidad oxidativa de las células de grasa parda es aproximadamente 20 veces mayor que la de las células de grasa blanca y casi 2 veces mayor que la capacidad oxidativa del músculo cardíaco. Cuando la temperatura ambiente disminuye, aumenta la actividad de los procesos oxidativos en el tejido adiposo pardo. En

Arroz. 8.14. La estructura del tejido adiposo blanco (según Yu. I. Afanasyev): A b- estructura ultramicroscópica de los adipocitos: 1 - núcleo de adipocitos; 2 - grandes gotas de lípidos; 3 - fibras nerviosas; 4 - hemocapilares; 5 - mitocondrias de adipocitos; V- tejido adiposo: 1 - adipocitos; 2 - vaso sanguíneo. Micrografía (tinción de Sudán III)

Arroz. 8.15. La estructura del tejido adiposo marrón (según Yu. I. Afanasyev): A- adipocitos de los que se ha eliminado la grasa mediante un microscopio óptico óptico; b- estructura ultramicroscópica de los adipocitos: 1 - núcleo de adipocitos; 2 - lípidos finamente triturados; 3 - numerosas mitocondrias; 4 - hemocapilares; 5 - fibra nerviosa
Esto libera energía térmica, que calienta la sangre en los capilares sanguíneos. El sistema simpático juega un papel determinado en la regulación del intercambio de calor. sistema nervioso y hormonas de la médula suprarrenal: adrenalina y norepinefrina, que estimulan la actividad de la lipasa tisular, que descompone los triglicéridos en glicerol y ácido graso. Esto conduce a la liberación de energía térmica, que calienta la sangre que fluye por los numerosos capilares entre los adipocitos. Durante el ayuno, el tejido adiposo marrón cambia menos que el tejido adiposo blanco.
Tejido mucoso
Tejido mucoso (texto mucoso) normalmente ocurre sólo en el embrión. El objeto clásico para su estudio es el cordón umbilical del feto humano (fig. 8.16). Los elementos celulares aquí están representados por un grupo heterogéneo de células que se diferencian de manera divergente de las células mesenquimales durante el período prenatal. Entre ellos se distinguen grandes fibroblastos en forma de estrella, miofibroblastos y células de músculo liso. Se distinguen por su capacidad para sintetizar vimentina, desmina, actina y miosina. Fibroblastos del tejido mucoso del canal umbilical.

Arroz. 8.16. Tejido conectivo mucoso del cordón umbilical: 1 - mucocitos; 2 - sustancia intercelular; 3 - pared de un vaso sanguíneo
La teca (“gelatina de Wharton”) sintetiza colágeno tipo IV, característico de las membranas basales, laminina y sulfato de heparina.
En la primera mitad del embarazo, el ácido hialurónico se encuentra en grandes cantidades entre las células de este tejido, lo que provoca la consistencia gelatinosa de la sustancia principal. Los fibroblastos de tejido conectivo gelatinoso sintetizan débilmente proteínas fibrilares. Sólo en las últimas etapas del desarrollo embrionario aparecen fibrillas de colágeno laxas en la sustancia gelatinosa.
8.2. TEJIDO ESQUELÉTICO
Tejidos esqueléticos (esqueletos de texto)- Estos son tipos de tejido conectivo con una función mecánica de soporte significativamente pronunciada, debido a la presencia de una sustancia intercelular densa: cartílago, tejido óseo, dentina y cemento dental. Además de su función principal, estos tejidos participan en el metabolismo agua-sal.
8.2.1. tejido cartilaginoso
tejido cartilaginoso (texto cartilaginoso) son parte de los órganos del sistema respiratorio, articulaciones, discos intervertebrales y otros, están formados por células - condrocitos Y condroblastos y una gran cantidad de sustancia hidrófila intercelular, caracterizada por su elasticidad. A esto se asocia la función de soporte del tejido cartilaginoso. El tejido cartilaginoso fresco contiene aproximadamente entre un 70 y un 80 % de agua, entre un 10 y un 15 % de sustancias orgánicas y entre un 4 y un 7 % de sales. Del 50 al 70% de la materia seca del tejido cartilaginoso es colágeno. El tejido del cartílago en sí no tiene vasos sanguíneos y los nutrientes se difunden desde el tejido circundante. pericondrio.
Clasificación. Hay tres tipos de tejido cartilaginoso: hialino, elástico, fibroso. Esta división del tejido del cartílago se basa en las características estructurales y funcionales de la estructura de su sustancia intercelular, el grado de contenido y la proporción de fibras colágenas y elásticas.
Desarrollo del tejido cartilaginoso (condrogistogénesis)
La fuente de desarrollo del tejido cartilaginoso es el mesénquima. Inicialmente, en algunas partes del cuerpo del embrión, donde se forma el cartílago, aparecen células mesenquimales.
perdemos nuestros procesos, nos multiplicamos vigorosamente y, muy adyacentes entre sí, creamos una cierta tensión: turgencia. Estas áreas se llaman rudimentos condrogénicos, o islotes condrogénicos(Figura 8.17). Contienen células madre que se diferencian en células condrogénicas semi-madre. Las células madre se caracterizan por una forma redonda, un alto valor de proporciones nuclear-citoplasmáticas, una disposición difusa de la cromatina y un pequeño nucléolo. Los orgánulos de estas células están poco desarrollados. En las células semi-madre (precondroblastos), aumenta el número de ribosomas libres, aparecen membranas del retículo endoplasmático de tipo granular, se alarga la forma de las células y disminuyen las relaciones núcleo-citoplasma. Posteriormente, los precursores poco diferenciados se diferencian en condroblastos morfológicamente identificables (v. fig. 8.17).
Condroblastos(del griego condros- cartílago, blastos- rudimento) son células jóvenes aplanadas capaces de proliferar y sintetizar la sustancia intercelular del cartílago (proteoglicanos). El citoplasma de los condroblastos tiene un retículo endoplásmico granular y agranular bien desarrollado, el complejo de Golgi. Cuando se tiñe, el citoplasma de los condroblastos resulta basófilo debido al rico contenido.
Arroz. 8.17. Desarrollo del tejido del cartílago hialino (esquema según Yu. I. Afanasyev): A- islote condrogénico; b- tejido cartilaginoso primario; V- etapas de diferenciación de condrocitos. 1 - células mesenquimales; 2 - células que se dividen mitóticamente; 3 - sustancia intercelular; 4 - sustancia intercelular oxifílica; 5 - sustancia intercelular basófila; 6 - condrocitos jóvenes; 7 - hemocapilar

Arroz. 8.18. Grupos isogénicos de células cartilaginosas:
1 - condrocito; 2 - matriz del territorio celular, que consta de una red en bucle de proteínas y proteoglicanos no colágenos; 3 - fibras de colágeno que forman la pared de la laguna; 4 - sección interterritorial de cartílago; 5 - proteoglicanos en la matriz interterritorial (según V.N. Pavlova, con modificaciones)
ARN. Los condroblastos sintetizan y secretan proteínas fibrilares (colágeno), aparece una sustancia intercelular que se tiñe de oxifílica. Así es como se forma el tejido cartilaginoso primario. Con la participación de condroblastos se produce. crecimiento periférico (aposicional) cartílago. Con una mayor diferenciación del tejido cartilaginoso, los condroblastos se convierten en condrocitos.
Condrocitos- el tipo principal de células del tejido cartilaginoso. Tienen forma ovalada, redonda o poligonal, según el grado de diferenciación. Están ubicados en cavidades especiales (lagunas) en la sustancia intercelular, solos o en grupos. Los grupos de células que se encuentran en una cavidad común se denominan isogénico(del griego isos- igual, génesis- desarrollo). Se forman por división de una célula (fig. 8.18). En los grupos isogénicos se distinguen tres tipos de condrocitos.
El primer tipo de condrocitos se caracteriza por una alta proporción núcleo-citoplasma, el desarrollo de elementos vacuolares del complejo de Golgi y la presencia de mitocondrias y ribosomas libres en el citoplasma. En estas células se observan a menudo figuras de división mitótica, lo que permite considerarlas como la fuente de aparición de grupos isogénicos de células (fig. 8.19). Los condrocitos tipo I predominan en el cartílago joven y en desarrollo. Los condrocitos tipo II se caracterizan por una disminución en la proporción núcleo-citoplasma, síntesis de ADN debilitada, preservación de un alto nivel de ARN, desarrollo intensivo del retículo endoplásmico granular y todos los componentes del complejo de Golgi, que aseguran la formación y secreción de glucosaminoglicanos. y proteoglicanos en la sustancia intercelular. Los condrocitos tipo III se caracterizan por la proporción núcleo-citoplasma más baja, un fuerte desarrollo y una disposición ordenada del retículo endoplásmico granular. Estas células conservan la capacidad de formar y secretar proteínas, pero su síntesis de glucosaminoglucanos se reduce.
A medida que el anillo cartilaginoso se desarrolla en la periferia, en el borde con el mesénquima, pericondrio- una capa que cubre el cartílago en desarrollo desde el exterior y que consta de una capa fibrosa externa y una capa condrogénica (cambial) interna. En la zona condrogénica, las células se dividen y diferencian intensamente en condroblastos, que conservan la capacidad de sintetizar ADN, reproducirse y también sintetizar componentes de la sustancia intercelular (colágeno de tipos I y III). En el proceso de secreción de productos de síntesis y colocación de capas sobre el cartílago existente a lo largo de su periferia, ellos mismos
las células están "amuralladas" en los productos de su actividad. Así crece el cartílago por superposición, o crecimiento aposicional.
Las células del cartílago que se encuentran en el centro del cartílago joven en desarrollo conservan la capacidad de dividirse mitóticamente durante algún tiempo, permaneciendo en la misma laguna (grupos isogénicos de células) y producen colágeno tipo II. Debido al aumento en el número de estas células y su producción de sustancia intercelular, aumenta la masa de cartílago desde el interior, lo que se llama crecimiento intersticial. El crecimiento intersticial se observa en la embriogénesis, así como durante la regeneración del tejido cartilaginoso.
Con el crecimiento y desarrollo del cartílago, sus secciones centrales se alejan cada vez más de los vasos cercanos y comienzan a experimentar dificultades en la nutrición, que se realiza de forma difusa desde los vasos del pericondrio. Como resultado, los condrocitos pierden su capacidad de reproducirse, algunos de ellos se destruyen y los proteoglicanos se convierten en una proteína oxífila más simple: el albumoide.
Por tanto, el diferencial condroblástico en la histogénesis embrionaria está representado por todas las formas de células en maduración. Sin embargo, posteriormente incluye sólo los enlaces medio y final (condroblastos y condrocitos) y es el único en los tejidos cartilaginosos.

Arroz. 8.19. Tres tipos de condrocitos (según Yu. I. Afanasyev):
A- Tipo i; b - Tipo II; V - III tipo. 1 - célula en división mitótica; 2 - retículo endoplásmico; 3 - mitocondrias; 4 - glucógeno; 5 - sustancia intercelular
Tejido de cartílago hialino
Tejido de cartílago hialino (texto cartilaginoso hialino), También llamado vítreo (del griego. hialos- vidrio) - debido a su transparencia y color blanco azulado, es la variedad más común

Arroz. 8.20. Cartílago hialino: A- microfotografía del cartílago hialino de la tráquea: 1 - pericondrio; 2 - condrocitos jóvenes; 3 - sustancia básica con grupos isogénicos de condrocitos ubicados en su interior (4)
densidad del tejido cartilaginoso. En el cuerpo adulto, el tejido hialino se encuentra en la unión de las costillas con el esternón, en la laringe, las vías respiratorias y en las superficies articulares de los huesos.
El tejido del cartílago hialino de varios órganos tiene mucho en común, pero al mismo tiempo difiere en la especificidad del órgano: la ubicación de las células, la estructura de la sustancia intercelular. La mayor parte del tejido del cartílago hialino que se encuentra en el cuerpo humano está cubierto pericondrio y representa formaciones anatómicas - cartílago.
El pericondrio tiene dos capas: la capa exterior, formada por tejido conectivo fibroso con vasos sanguíneos; interno, predominantemente celular, que contiene condroblastos y sus precursores: precondroblastos. Debajo del pericondrio, en la capa superficial, se encuentran condrocitos jóvenes en forma de huso, cuyo eje longitudinal se dirige a lo largo de la superficie del cartílago (fig. 8.20). En las capas más profundas, las células del cartílago adquieren una forma ovalada o redonda. Debido al hecho de que los procesos sintéticos y secretores en estas células están debilitados, después de la división no divergen mucho, sino que se encuentran de manera compacta, formando los llamados grupos isógenos de 2 a 4 condrocitos.
Las células cartilaginosas más diferenciadas y los grupos isogénicos, además de la capa pericelular oxifílica de la sustancia intercelular, tienen una zona basófila ubicada hacia afuera. Estas propiedades se explican por la distribución desigual de los componentes químicos de la sustancia intercelular: proteínas y glucosaminoglicanos.
En el cartílago hialino de cualquier localización se acostumbra distinguir áreas territoriales sustancia intercelular o matriz (ver Fig. 8.18). La región territorial incluye la matriz que rodea inmediatamente a las células cartilaginosas o sus grupos. En estas áreas, las fibras y fibrillas de colágeno tipo II, al retorcerse, rodean grupos isogénicos de células cartilaginosas, protegiéndolas de la presión mecánica. en interterritorial

Arroz. 8.20. Continuación
b- diagrama de la estructura del cartílago y pericondrio (Fig. Yu. I. Afanasyev): 1 - capa fibrosa exterior; 2 - capa celular interna; 3 - tejido cartilaginoso; V- componentes celulares y fibrosos del cartílago articular (según V.P. Modyaev, V.N. Pavlova, con modificaciones). I - zona superficial; II - zona intermedia; III - zona basal (profunda); IV - hueso subcondral; A - componentes celulares del cartílago articular: 1 - placa acelular; 2 - condrocitos de la capa tangencial; 3 - condrocitos de la región de transición; 4 - grupos isogénicos; 5 - “columnas” de condrocitos; 6 - condrocitos hipertrofiados; 7 - línea basófila (límite) entre cartílago calcificado y no calcificado; 8 - cartílago calcificante; B - sistema fibrilar del cartílago articular: 1 - placa acelular; 2 - fibras tangenciales de la zona superficial; 3 - direcciones principales de las fibras de colágeno en la zona intermedia; 4 - fibras radiales de la capa basal; 5 - línea basófila (límite)
En esta matriz, las fibras de colágeno están orientadas en la dirección del vector de acción de las fuerzas de las cargas principales. El espacio entre las estructuras de colágeno está lleno de proteoglicanos.
La condronectina juega un papel importante en la organización estructural de la sustancia intercelular del cartílago. Esta glicoproteína conecta las células entre sí y con diversos sustratos (colágeno, glucosaminoglicanos). La función biomecánica de soporte del tejido cartilaginoso durante la compresión y la tensión está garantizada no solo por la estructura de su marco fibroso, sino también por la presencia de proteoglicanos hidrófilos con un alto nivel de hidratación (65-85%). La alta hidrofilia de la sustancia intercelular favorece la difusión de nutrientes y sales. Los gases y muchos metabolitos también difunden libremente a través de él. Sin embargo, las moléculas de proteínas grandes con propiedades antigénicas no pasan. Esto explica el trasplante exitoso de secciones de cartílago en la clínica (trasplante de una persona a otra). El metabolismo de los condrocitos es predominantemente anaeróbico, glucolítico.
Sin embargo, no todo el cartílago tiene la misma estructura. Una característica estructural del cartílago hialino de la superficie articular es la ausencia de pericondrio en la superficie que mira hacia la cavidad articular. El cartílago articular consta de tres zonas mal delimitadas: superficial, intermedia y basal (ver Fig. 8.20, c).
En la zona superficial del cartílago articular hay condrocitos pequeños, aplanados y no especializados, que en su estructura se asemejan a los fibrocitos.
En la zona intermedia, las células son más grandes, de forma redonda, metabólicamente muy activas: con grandes mitocondrias, un retículo endoplásmico granular bien desarrollado y un complejo de Golgi con numerosas vesículas.
La zona profunda (basal) está dividida por una línea basófila en capas calcificantes y no calcificantes. Este último es atravesado por vasos sanguíneos del hueso subcondral subyacente. Una característica de la sustancia intercelular de la zona profunda del cartílago articular es el contenido de vesículas de matriz densa, estructuras de membrana con un diámetro de 30 nm a 1 μm, que son los lugares de mineralización inicial de los tejidos esqueléticos (además del cartílago, se encuentran en el tejido óseo y la predentina). Las estructuras de membrana se forman por abultamiento de una sección del plasmalema de condrocitos (respectivamente, osteoblasto en tejido óseo y odontoblasto en predentina), seguido de gemación desde la superficie celular y distribución localizada en zonas de mineralización. También pueden ser producto de la desintegración celular completa. El cartílago articular se nutre sólo parcialmente de los vasos de la zona profunda, pero principalmente del líquido sinovial de la cavidad articular.
Tejido cartilaginoso elástico
Tejido cartilaginoso elástico (texto cartilaginoso elástico) Se encuentra en aquellos órganos donde la base cartilaginosa está sujeta a flexión (en el pabellón auricular, cartílagos corniculados y en forma de cuña de la laringe, etc.). En estado fresco y no fijado, el tejido cartilaginoso elástico es de color amarillento y no tan transparente como el tejido hialino. Según la estructura general del cartílago elástico, es similar al cartílago hialino. Por fuera está cubierto de pericondrio. Células cartilaginosas (jóvenes y especializadas). condrocitos) se encuentran individualmente en cápsulas o forman grupos isogénicos. Una de las principales diferencias
Una característica importante del cartílago elástico es la presencia en su sustancia intercelular, junto con las fibras de colágeno, de fibras elásticas que penetran la sustancia intercelular en todas las direcciones (fig. 8.21).
Desde la capa adyacente al pericondrio, las fibras elásticas pasan sin interrupción a las fibras elásticas del pericondrio. Hay menos lípidos, glucógeno y sulfatos de condroitina en el cartílago elástico que en el cartílago hialino.

Arroz. 8.21. Tejido cartilaginoso elástico. Microfotografía teñida con orceína: 1 - grupos isogénicos de condrocitos; 2 - fibras elásticas
Tejido cartilaginoso fibroso
Tejido cartilaginoso fibroso (texto cartilaginoso fibroso) ubicado en los discos intervertebrales, articulaciones semimóviles, en los lugares de transición del tejido conectivo fibroso (tendones, ligamentos) al cartílago hialino, donde los movimientos limitados van acompañados de fuertes tensiones. La sustancia intercelular contiene haces de colágeno dirigidos en paralelo,
aflojándose gradualmente y convirtiéndose en cartílago hialino. El cartílago tiene cavidades en las que se encierran las células del cartílago. Estos últimos se encuentran solos o forman pequeños grupos isogénicos. El citoplasma de las células suele estar vacuolado. Desde el cartílago hialino hacia el tendón, el fibrocartílago se vuelve cada vez más parecido a un tendón. En el borde del cartílago y el tendón, entre los haces de colágeno, hay columnas de células de cartílago comprimidas que, sin ningún borde, pasan a las células del tendón ubicadas en el tejido conectivo denso (fig. 8.22).
Cambios relacionados con la edad. A medida que el cuerpo envejece, la concentración de proteoglicanos y la hidrofilia asociada en el tejido del cartílago disminuyen. Los procesos de reproducción de condroblastos y condrocitos jóvenes están debilitados. En el citoplasma de estas células, el volumen del complejo de Golgi, el retículo endoplásmico granular y las mitocondrias disminuye y la actividad de las enzimas disminuye.
Participar en la reabsorción de células y sustancias intercelulares modificadas distróficamente. condroclastos, Morfológicamente idéntico a los osteoclastos. Después de la muerte de los condrocitos, algunos de los huecos se llenan de sustancia amorfa y fibrillas de colágeno. En lugares del intercelular

Arroz. 8.22. Tejido cartilaginoso fibroso. Sección del disco intervertebral: 1 - fibras de colágeno; 2 - condrocitos
En la sustancia se detectan depósitos de sales de calcio (“tizaje del cartílago”), como resultado de lo cual el cartílago se vuelve turbio, duro y quebradizo. Como resultado, la alteración resultante en el trofismo de las zonas centrales del cartílago puede provocar el crecimiento de vasos sanguíneos en ellas, seguido de la formación de hueso.
Regeneración. La regeneración fisiológica del tejido cartilaginoso se lleva a cabo gracias a las células poco diferenciadas del pericondrio y del cartílago, mediante la reproducción y diferenciación de precondroblastos y condroblastos. Sin embargo, este proceso es muy lento. La regeneración postraumática del tejido cartilaginoso de localización extraarticular se lleva a cabo gracias al pericondrio (fig. 8.23).
En el cartílago articular, dependiendo de la profundidad de la lesión, la regeneración se produce tanto debido a la proliferación de células únicas en grupos isogénicos (con daño superficial) como debido a la segunda fuente de regeneración: las células cambiales del tejido óseo subcondral (con daño profundo a cartílago), que forman la matriz orgánica del hueso - osteoide.
En cualquier caso, los procesos distróficos (necróticos) se observan directamente en el área de la lesión del tejido del cartílago y los condroblastos en proliferación se ubican de manera más centrífuga. En la herida se forma tejido conectivo fibroso, que posteriormente es reemplazado por cartílago. En general, la regeneración se completa entre 3 y 6 meses después de la lesión.
Factores que regulan el metabolismo del tejido cartilaginoso. La regulación del metabolismo del tejido cartilaginoso se produce bajo la influencia de cargas mecánicas, factores nerviosos y hormonales. La presión periódica sobre el tejido cartilaginoso y el debilitamiento de la carga son factores que actúan constantemente para la difusión de nutrientes disueltos en agua, productos metabólicos y reguladores hormonales-humorales de los capilares del pericondrio, que tiene receptores y efectores, o del líquido sinovial de las articulaciones. . Además, los condrocitos tienen citorreceptores para varias hormonas que circulan en la sangre. Entonces, las hormonas pituitarias son somatotropina.
y prolactina: estimulan el crecimiento del tejido del cartílago, pero no afectan su maduración. Las hormonas tiroideas (tiroxina y triyodotironina) aceleran la citodiferenciación de los condrocitos, pero inhiben los procesos de crecimiento en el cartílago. Las hormonas de las glándulas tiroides y paratiroides (calcitonina y hormona paratiroidea) tienen un efecto similar sobre el metabolismo del cartílago y ayudan a estimular los procesos de crecimiento, pero en menor medida su maduración. La hormona de los islotes endocrinos del páncreas, la insulina, mejora la citodiferenciación de las células mesenquimales esqueléticas y, en las etapas de la ontogénesis posnatal, tiene un efecto mitogénico y de crecimiento. Las hormonas de la corteza suprarrenal (glucocorticoides y la hormona sexual femenina estrógeno) inhiben la biosíntesis de colágeno y glucosaminoglucanos en los condrocitos y, en el período posnatal temprano, sus altas concentraciones contribuyen al envejecimiento del tejido del cartílago y a cambios destructivos en el mismo. La hormona sexual masculina, la testosterona, estimula la biosíntesis de glicosaminoglicanos no sulfatados, lo que conduce a una disminución en la maduración del tejido cartilaginoso. En general, cabe señalar que las hormonas regulan procesos metabólicos específicos en los condrocitos, pero la capacidad de los condrocitos para responder a su acción depende tanto del estado endocrino del organismo (normal, deficiente o excesivo de hormonas) como de la Estado estructural y funcional de los propios condrocitos.
8.2.2. Tejido óseo
Tejido óseo (textus ossei) Es un tipo polidiferente especializado de tejido conectivo con alta mineralización del tejido intercelular.

Arroz. 8.23. Regeneración postraumática del cartílago hialino con poca profundidad. (un 1 Y
b 1 y profundo (un 2 y b 2) daño: A\ 2 - regeneración del cartílago articular: 1 - membrana sinovial; 2 - cartílago; 3 - hueso; 4 - zona de necrosis; 5 - zona de proliferación; 6 - cartílago no calcificado; 7 - cartílago calcificado; 8 - osteonas con vasos; 9 - médula ósea; 10 - tejido de granulación; b - regeneración del cartílago costal: 1 - pericondrio; 2 - cartílago; 3 - zona de necrosis; 4 - zona de proliferación; 5 - tejido de granulación (según V.N. Pavlova). Flechas: direcciones de movimiento de células y tejidos en el proceso de la herida.
Materia orgánica que contiene alrededor del 70% de compuestos inorgánicos, principalmente fosfatos de calcio. Más de 30 microelementos (cobre, estroncio, zinc, bario, magnesio, etc.) que intervienen en papel vital en los procesos metabólicos del cuerpo.
Materia orgánica - matriz tejido óseo: representado principalmente por proteínas y lípidos de tipo colágeno. En comparación con la matriz del tejido cartilaginoso, contiene una cantidad relativamente pequeña de agua, ácido condroitinsulfúrico, pero muchos ácidos cítrico y otros que forman complejos con el calcio, impregnando la matriz orgánica del hueso. Los componentes orgánicos e inorgánicos en combinación entre sí determinan las propiedades mecánicas: la capacidad de resistir el estiramiento, la compresión, etc. De todos los tipos de tejidos conectivos, las funciones de soporte, mecánicas y protectoras son más pronunciadas en el tejido óseo. Para los órganos internos, también es un depósito de sales de calcio, fósforo, etc.
A pesar del alto grado de mineralización, los tejidos óseos sufren una renovación constante de sus sustancias constituyentes, una destrucción y creación constantes y una adaptación debido a las condiciones cambiantes de funcionamiento. Las propiedades morfofuncionales del tejido óseo cambian según la edad, la actividad muscular, las condiciones nutricionales, así como bajo la influencia de la actividad de las glándulas endocrinas, la inervación, etc.
Clasificación. Hay dos tipos principales de tejido óseo: fibroso grueso (malla fibrosa) Y laminar. Estos tipos de tejido óseo difieren en estructura y propiedades físicas, que están determinados principalmente por la estructura de la sustancia intercelular. Los tejidos óseos también incluyen dentina Y cemento dientes que son similares al tejido óseo en términos del alto grado de mineralización de la sustancia intercelular (ver Capítulo 16) y función mecánica de soporte.
Tejido óseo fibroso rugoso
Tejido óseo fibroso rugoso (texto óseo reticulofibroso) Ocurre principalmente en embriones. En los adultos, se puede encontrar en el sitio de suturas craneales demasiado crecidas, en los sitios de unión de los tendones a los huesos. Las fibras de colágeno dispuestas aleatoriamente forman en él haces gruesos, claramente visibles incluso con aumentos microscópicos relativamente bajos (fig. 8.24).
En la sustancia principal del tejido óseo de fibras gruesas hay formas ovaladas alargadas. cavidades óseas, o lagunas, con largos túbulos anastomosados en los que se encuentran las células óseas - osteocitos con sus brotes. La superficie del hueso está cubierta. periostio.
Tejido óseo laminar
Tejido óseo laminar (texto óseo laminar)- el tipo de tejido óseo más común en el cuerpo adulto. Consiste en placas óseas (lamellae ossea). Grosor y longitud de este último.

Arroz. 8.24. La estructura del tejido óseo de fibras gruesas (según Yu. I. Afanasyev): 1 - haces de fibras de colágeno entretejidas; 2 - osteocitos
varía de varias decenas a cientos de micrómetros, respectivamente. No son monolíticos, sino que contienen fibrillas de colágeno (oseína) dirigidas paralelamente, orientadas en diferentes planos. En la parte central de las placas, las fibrillas tienen una dirección predominantemente longitudinal; a lo largo de la periferia se añaden direcciones tangenciales y transversales. Las láminas pueden deslaminarse y las fibrillas de una lámina pueden continuar hacia las adyacentes, creando una única base fibrosa del hueso. Además, las placas óseas están atravesadas por fibrillas y fibras individuales, orientadas perpendicularmente a las placas óseas, entretejidas en las capas intermedias entre ellas, logrando así una mayor resistencia del tejido óseo laminar (fig. 8.25). La sustancia compacta y esponjosa de la mayoría de los huesos planos y tubulares del esqueleto se forma a partir de este tejido.
Desarrollo del tejido óseo (osteohistogénesis)
El desarrollo del tejido óseo en el embrión se realiza de dos formas: 1) directamente desde el mesénquima (osteogénesis directa); 2) del mesénquima en lugar de un modelo óseo cartilaginoso previamente desarrollado (osteogénesis indirecta). El desarrollo postembrionario del tejido óseo ocurre durante la regeneración fisiológica y reparadora.
El tejido óseo incluye osteoblástico Y diferenciales osteoclásticos. La primera (principal) consta de una serie de células diferenciadoras: células madre, células semi-madre (preosteoblastos), osteoblastos (un tipo

Arroz. 8.25. La estructura del tejido óseo laminar (según Yu. I. Afanasyev):
1 - placas óseas; 2 - osteocitos; 3 - contactos de procesos de osteocitos; 4 -
Fibras de colágeno orientadas dentro de cada placa ósea.
paralelo
fibroblastos), osteocitos. Los procesos de diferenciación osteogénica de las células están influenciados por factores osteogénicos (proteína morfogenética ósea), presión parcial de oxígeno en el tejido, presencia de fosfatasa alcalina, etc. El segundo diferencial (asociado) incluye osteoclastos(un tipo de macrófago) que se desarrolla a partir de células madre sanguíneas.
Las células osteogénicas madre y semitallo no se identifican morfológicamente.
Osteoblastos(del griego osteona- hueso, blastos- rudimento) son células jóvenes que crean tejido óseo. En el hueso formado, se encuentran solo en las capas profundas del periostio y en los lugares de regeneración del tejido óseo después de una lesión. Son capaces de proliferar; en el hueso en formación cubren toda la superficie del haz de hueso en desarrollo con una capa casi continua (fig. 8.26). La forma de los osteoblastos puede ser diferente: cúbica, piramidal o angular. El tamaño de su cuerpo es de unas 15-20 micras. El núcleo es de forma redonda u ovalada, a menudo ubicado de forma excéntrica y contiene uno o más nucléolos.

Arroz. 8.26. Osteogénesis "directa":
A- isla osteogénica (diagrama); b- etapa osteoide (esquema); V- osificación de la sustancia intercelular (esquema); GRAMO- Osteogénesis “directa” en hueso plano (micrografía). 1 - células mesenquimales; 2 - capilares sanguíneos; 3 - osteoblastos; 4 - osteoide; 5 - sustancia intercelular mineralizada; 6 - osteocito; 7 - osteoclasto
En el citoplasma de los osteoblastos, el retículo endoplásmico granular, las mitocondrias y el complejo de Golgi están bien desarrollados (fig. 8.27). En él se detectan cantidades importantes de ARN y una alta actividad de fosfatasa alcalina. Los osteoblastos secretan vesículas de matriz que contienen lípidos.

Arroz. 8.27. La estructura de un osteoblasto (según Yu. I. Afanasyev):
a - a nivel óptico de luz; b- a nivel ultramicroscópico. 1 - núcleo; 2 - citoplasma; 3 - retículo endoplásmico granular; 4 - osteoide; 5 - tejido óseo mineralizado
Ca 2+, fosfatasa alcalina, que conduce a la calcificación de la matriz del tejido orgánico.
osteocitos(del griego osteona- hueso, cito- célula) es el número predominante de células definitivas del tejido óseo que han perdido la capacidad de dividirse. Tienen una forma ramificada, un núcleo compacto y relativamente grande y un citoplasma ligeramente basófilo (fig. 8.28). Los orgánulos están poco desarrollados. No se ha establecido la presencia de centríolos en los osteocitos.
Las células óseas se encuentran en cavidades o lagunas óseas que siguen los contornos del osteocito. La longitud de las cavidades varía de 22 a 55 micrones, el ancho, de 6 a 14 micrones. Los túbulos de las cavidades óseas están llenos de líquido tisular, se anastomosan entre sí y con el peri-
espacios esculares de los vasos que entran en el hueso. El intercambio de sustancias entre los osteocitos y la sangre se produce a través del líquido tisular.
osteoclastos(del griego osteona- hueso y clastos- fragmentado). Estas células de naturaleza hematógena son capaces de destruir el cartílago y el hueso calcificados. Su diámetro alcanza las 150-180 micrones y contienen de 3 a varias docenas de núcleos (fig. 8.29). El citoplasma es ligeramente basófilo, a veces oxifílico. Los osteoclastos suelen localizarse en la superficie de las trabéculas óseas. En el lado del osteoclasto adyacente a la superficie destruida, hay un borde microplegado (corrugado); es el área de síntesis y secreción de enzimas hidrolíticas. A lo largo de la periferia del osteoclasto hay una zona de estrecha adherencia de la célula a la superficie del hueso, que, por así decirlo, sella el área de acción de las enzimas. Esta zona del citoplasma es clara y contiene pocos orgánulos, a excepción de los microfilamentos de actina.
La capa periférica de citoplasma sobre el borde corrugado contiene numerosas vesículas pequeñas y vacuolas más grandes. Se cree que los osteoclastos liberan CO 2 en ambiente, y la enzima hidrasa carbónica, que se encuentra aquí, promueve la formación de ácido (H 2 CO 3) y la disolución de compuestos de calcio. El osteoclasto es rico en mitocondrias y lisosomas, cuyas enzimas (colagenasa y otras proteasas) descomponen el colágeno y los proteoglicanos de la matriz del tejido óseo. En el lugar de contacto del osteoclasto con la sustancia ósea, se forma en esta última una laguna de resorción. Un osteoclasto puede destruir tanto hueso como crean 100 osteoblastos al mismo tiempo. Funciones de los osteoblastos
Los sts y los osteoclastos están interconectados y se correlacionan con la participación de hormonas, prostaglandinas, carga funcional, vitaminas, etc.
Sustancia intercelular (sustancia intercelular) Consiste en una sustancia básica amorfa impregnada de sales inorgánicas, en la que se ubican fibras de colágeno formando pequeños haces. Contienen (hasta un 90%) proteína: colágeno tipo I. Las fibras pueden tener una dirección aleatoria (en tejido óseo fibroso grueso) o estrictamente orientada (en tejido óseo laminar).
La sustancia fundamental del tejido óseo, en comparación con el cartílago, contiene una cantidad relativamente pequeña de ácido condroitinsulfúrico, pero una gran cantidad de ácidos cítrico y otros que forman complejos con el calcio, impregnando la matriz orgánica del hueso. Además de la proteína de colágeno, en la sustancia principal del tejido óseo se encuentran proteínas no colagenosas (osteocalcina, sialoproteína, osteonectina, osteopontina, etc., que participan en los procesos de mineralización), así como glucosaminoglicanos. La sustancia fundamental del hueso contiene cristales de hidroxiapatita, ordenados en relación con las fibrillas de la matriz orgánica del hueso, así como fosfato cálcico amorfo. En el tejido óseo se encuentran más de 30 oligoelementos (cobre, estroncio, zinc, bario, magnesio, etc.), que desempeñan un papel vital en los procesos metabólicos del organismo. Un aumento sistemático de la actividad física conduce a un aumento de la masa ósea del 10 al 50% debido a la alta mineralización.
Osteohistogénesis directa. Este método de osteogénesis es característico del desarrollo de tejido óseo de fibras gruesas durante la formación de planos.
Arroz. 8.28. La estructura de un osteocito (según Yu. I. Afanasyev):
A- a nivel óptico de luz; b- a nivel ultramicroscópico. 1 - procesos de osteocitos; 2 - núcleo; 3 - retículo endoplásmico; 4 - complejo de Golgi; 5 - mitocondrias; 6 - sustancia ósea osteoide (no calcificada) a lo largo del borde de la laguna en la que se encuentran los osteocitos

Arroz. 8.29. La estructura de un osteoclasto (según Yu. I. Afanasyev):
A- a nivel óptico de luz; b- a nivel ultramicroscópico. 1 - núcleo; 2 - borde ondulado del osteoclasto; 3 - zona de luz; 4 - lisosomas; 5 - zona de reabsorción de sustancia intercelular; 6 - sustancia mineralizada
huesos, como los huesos tegumentarios del cráneo. Este proceso se observa principalmente durante el primer mes de desarrollo intrauterino y se caracteriza por la formación de tejido óseo osteoide "membranoso" primario, seguida de la impregnación (deposición) de calcio, fósforo y otras sales en la sustancia intercelular. En la primera etapa- formación de una isla esquelética: en los lugares de desarrollo del futuro hueso, se produce una proliferación focal de células mesenquimales y una vascularización de la isla esquelética. Las células mesenquimales se orientan según los vectores de carga y se diferencian en precursores osteogénicos: los preosteoblastos. En la segunda etapa Las células de los islotes se diferencian en osteoblastos, en los que comienza la biosíntesis de proteínas de colágeno y su secreción, lo que da como resultado la aparición de una sustancia intercelular oxifílica con fibrillas de colágeno, la matriz orgánica del tejido óseo (etapa osteoide). Las fibras en crecimiento separan las células, que permanecen conectadas entre sí mediante procesos en desarrollo. Así, los osteoblastos adquieren una forma procesada y se convierten en osteocitos incluidos en el espesor de la masa fibrosa, y pierden la capacidad de reproducirse. Las mucoproteínas (oseomucoide) aparecen en la sustancia principal, cementando las fibras en una masa fuerte. Al mismo tiempo, a partir del mesénquima circundante se forman nuevas generaciones de osteoblastos, en los que crece hueso desde el exterior (crecimiento aposicional).
Tercera etapa- calcificación (impregnación con sales) de la sustancia intercelular. En este caso, los osteoblastos secretan la enzima fosfatasa alcalina, que descompone los glicerofosfatos contenidos en la sangre periférica en compuestos de carbohidratos (azúcares) y ácido fosfórico. Este último reacciona con las sales de calcio, que se depositan en la sustancia principal y las fibras, primero en forma de compuestos de calcio que forman amorfos.
A partir de él se forman depósitos de [Ca 3 (PO 4) 2], posteriormente se forman cristales de hidroxiapatita [Ca 10 (PO 4) 6 (OH) 2].
Uno de los mediadores de la calcificación es osteonectina- una glicoproteína que une selectivamente las sales de calcio y fósforo al colágeno. Como resultado de la calcificación, se forman barras o vigas transversales de hueso. Luego, de estas barras transversales se ramifican excrecencias, interconectadas y formando una amplia red. Los espacios entre las barras transversales están ocupados por tejido conectivo fibroso laxo a través del cual pasan vasos sanguíneos.
Cuando se completa la histogénesis, aparece una gran cantidad de fibras y células osteogénicas en el tejido conectivo embrionario a lo largo de la periferia del germen óseo. Parte de este tejido conectivo fibroso adyacente directamente a las trabéculas óseas se convierte en periostio, lo que proporciona trofismo y regeneración ósea. Este hueso, que aparece en las etapas del desarrollo embrionario y está formado por puntales de tejido óseo fibroso grueso, se denomina hueso esponjoso primario. En etapas posteriores de desarrollo, es reemplazado por hueso esponjoso secundario de los adultos, que se diferencia del primero en que está formado a partir de tejido óseo laminar (la cuarta etapa de la osteogénesis). El desarrollo del tejido óseo laminar está estrechamente relacionado con el proceso de destrucción de secciones individuales de hueso y el crecimiento de vasos sanguíneos hacia el interior del espesor del hueso de fibras gruesas. Los osteoclastos participan en este proceso tanto durante la osteogénesis embrionaria como después del nacimiento (v. fig. 8.29). Se forma una capa de osteoblastos alrededor de los vasos sanguíneos y posteriormente aparecen nuevas placas de osteonas concéntricas. Las fibras de colágeno de cada lámina son paralelas, pero orientadas en ángulo con respecto a las fibras de la lámina anterior. Así, se forman cilindros óseos alrededor del vaso, insertados uno dentro del otro (osteones primarios). Desde el momento en que aparecen las osteonas, el tejido óseo fibroso grueso deja de desarrollarse y es sustituido por tejido óseo laminar. Desde el lado del periostio, se forman placas circundantes externas (generales, generales), que cubren todo el hueso desde el exterior. Así se desarrollan los huesos planos. Posteriormente, el hueso formado en el período embrionario sufre una reestructuración: las osteonas primarias se destruyen y se desarrollan nuevas generaciones de osteonas. Esta reestructuración ósea prácticamente continúa durante toda la vida.
A diferencia del tejido cartilaginoso, el hueso siempre crece aplicando tejido nuevo al tejido existente, es decir, mediante aposiciones, y el suministro de sangre óptimo es necesario para la diferenciación de las células esqueléticas de los islotes.
Osteohistogénesis indirecta. En el segundo mes de desarrollo embrionario, en los lugares de los futuros huesos tubulares, se forma un rudimento cartilaginoso a partir del mesénquima, que muy rápidamente toma la forma del futuro hueso (modelo cartilaginoso). El rudimento consiste en cartílago hialino embrionario cubierto por pericondrio (fig. 8.30). Durante algún tiempo crece tanto por las células formadas en el lado del pericondrio como por la proliferación de células en las zonas internas.

Arroz. 8.30. Osteogénesis indirecta (cartilaginosa). Formación de un modelo cartilaginoso de hueso y manguito de hueso pericondral (según Yu. I. Afanasyev):
a-d - etapas de la osteogénesis. 1 - modelo cartilaginoso primario de un hueso tubular; 2 - pericondrio; 3 - tejido cartilaginoso; 4 - manguito de hueso pericondral; 5 - periostio; 6 - columnas de células cartilaginosas; 7 - zona de células vesiculares; 8 - mesénquima que crece hacia el cartílago con osteoclastos diferenciados (9) y capilares sanguíneos (10); 11 - osteoblastos; 12 - tejido óseo formado endocondral; 13 - punto de osificación en la epífisis
El desarrollo del hueso en lugar del cartílago, es decir, la osteogénesis indirecta, comienza en la zona de la diáfisis (osificación pericondral). Educación anillo óseo pericondral(manguito) está precedido por la proliferación de vasos sanguíneos con diferenciación en el pericondrio adyacente a la parte media de la diáfisis de los osteoblastos, que primero forman un manguito tejido óseo fibroso grueso(centro de osificación primario), luego reemplazado laminar.
La formación de un manguito óseo altera la nutrición del cartílago. Como resultado, se producen cambios distróficos en el centro de la parte diafisaria del rudimento cartilaginoso. Los condrocitos se vacuolan, sus núcleos se picnotizan y se forman los llamados condrocitos vesiculares. Se detiene el crecimiento del cartílago en este lugar. La expansión del anillo óseo pericondral se acompaña de un aumento en la zona de destrucción del cartílago y la aparición de osteoclastos, que despejan el camino para los vasos sanguíneos y los osteoblastos que crecen hacia el modelo óseo tubular (ver Fig. 8.30). Esto conduce a la aparición de focos de osificación endocondral (centros de osificación secundarios). Debido al crecimiento continuo de las secciones distales adyacentes sin cambios de la diáfisis, los condrocitos en el borde de la epífisis y la diáfisis se recogen en columnas, cuya dirección coincide con el eje longitudinal del futuro hueso. Por lo tanto, en la columna de condrocitos hay dos procesos dirigidos de manera opuesta: reproducción y crecimiento en las partes distales de la diáfisis y procesos distróficos en su parte proximal. Al mismo tiempo, se depositan sales minerales entre las células inflamadas, provocando la aparición de basofilia severa y fragilidad del cartílago.
A partir del momento en que crece la red vascular y aparecen los osteoblastos, el pericondrio se reconstruye convirtiéndose en periostio. Posteriormente, los vasos sanguíneos con el mesénquima circundante, las células osteogénicas y los osteoclastos crecen a través de las aberturas del manguito óseo y entran en contacto con el cartílago calcificado (ver Fig. 8.30). Bajo la influencia de enzimas secretadas por los osteoclastos, se produce la disolución (condrólisis) de la sustancia intercelular calcificada. El cartílago diafisario se destruye, aparecen espacios alargados en él, en los que los osteoblastos se "asientan", formando tejido óseo en la superficie de las áreas restantes de la sustancia intercelular calcificada del cartílago.
Centro de osificación primario o diafisario. El proceso de formación de hueso dentro del rudimento cartilaginoso se llama osificación endocondral (del griego. finalizará el- adentro).
Simultáneamente con el desarrollo del hueso endocondral, aparecen signos de su destrucción por parte de los osteoclastos. Debido a la destrucción del tejido óseo endocondral, se forman cavidades y espacios aún más grandes (cavidades de reabsorción) y, finalmente, una cavidad de la médula ósea. A partir del mesénquima que penetra aquí se forma el estroma de la médula ósea, en el que se asientan las células madre de la sangre y del tejido conectivo. Al mismo tiempo, aparecen cada vez más barras transversales nuevas de tejido óseo a lo largo de la periferia de la diáfisis desde el lado del periostio. El tejido óseo, que crece en longitud hacia las epífisis y aumenta en grosor, forma una densa capa de hueso.
La organización del hueso perióstico se produce de manera diferente a la organización del tejido óseo endocondral. En lugar del colapso del hueso fibroso grueso, alrededor de los vasos que han penetrado aquí, que discurren a lo largo del eje longitudinal del rudimento óseo, comienzan a formarse placas concéntricas, que consisten en finas fibras de colágeno orientadas paralelamente y una sustancia intercelular cementante. Así surgen osteonas primarias. Su luz es amplia, los límites de las placas no están muy definidos. Tras la aparición de la primera generación de osteonas, el desarrollo de placas envolventes externas (generales), rodeando el hueso en la zona de la diáfisis. Después de la diáfisis, aparecen centros de osificación en las epífisis. Esto va precedido primero de la diferenciación de los condrocitos, su hipertrofia, seguido de un deterioro de la nutrición, degeneración y calcificación de la sustancia intercelular. Posteriormente se observa un proceso de osificación similar al descrito anteriormente. La osificación se acompaña de un crecimiento hacia el interior de las epífisis de los vasos sanguíneos.
En la zona intermedia entre la diáfisis y las epífisis se conserva tejido cartilaginoso. cartílago metafisario, siendo una zona de crecimiento óseo en longitud.
Estructura histológica del hueso tubular como órgano.
El hueso tubular como órgano se construye principalmente a partir de tejido óseo laminar, excepto los tubérculos. La parte exterior del hueso está recubierta por periostio, a excepción de las superficies articulares de las epífisis, que están recubiertas por una especie de cartílago hialino.

Arroz. 8.31. La estructura del hueso tubular (según V. G. Eliseev, Yu. I. Afanasyev, E. F. Kotovsky):
A- periostio; b- sustancia ósea compacta; V- endosté; GRAMO- cavidad de la médula ósea. 1 - capa de placas comunes exteriores; 2 - osteona; 3 - canal de osteonas; 4 - placas de inserción; 5 - capa de placas comunes internas; 6 - trabécula ósea de sustancia esponjosa; 7 - capa fibrosa de periostio; 8 - vasos sanguíneos del periostio; 9 - canal de perforación; 10 - osteocitos
Periostio, o periostio. El periostio tiene dos capas: externa (fibrosa) e interna (celular). La capa exterior está formada principalmente por tejido conectivo fibroso (fig. 8.31, 8.32). La capa interna contiene una gran cantidad de células: células cambiales, preosteoblastos y osteoblastos de diversos grados de diferenciación. Las células cambiales fusiformes tienen un pequeño volumen de citoplasma y un aparato sintético moderadamente desarrollado. Los preosteoblastos son células de forma ovalada que proliferan vigorosamente y son capaces de sintetizar glucosaminoglicanos. Los osteoblastos se caracterizan por tener un aparato de síntesis de proteínas (colágeno) bien desarrollado. Los vasos y nervios que irrigan el hueso pasan a través del periostio.
El periostio conecta el hueso con los tejidos circundantes y participa en su trofismo, desarrollo, crecimiento y regeneración.
La estructura de la diáfisis. La sustancia compacta que forma la diáfisis ósea está formada por placas óseas cuyo espesor oscila entre 4 y 12-15 micrones. Las placas óseas están dispuestas en un orden determinado,

Arroz. 8.32. Periostio (según Yu. I. Afanasyev):
1 - capa exterior (fibrosa); 2 - capa interna (celular); 3 - células osteogénicas; 4 - tejido óseo
formando sistemas complejos. La diáfisis tiene tres capas: exterior capa de placas circundantes (generales, generales), promedio, formado por placas óseas colocadas concéntricamente alrededor de los vasos: osteonas y llamada capa de osteonas (fig. 8.33), y interior capa de placas circundantes (comunes).
Las placas exteriores no forman anillos completos alrededor de la diáfisis ósea; están superpuestas en la superficie por capas de placas posteriores. Las placas internas están bien desarrolladas sólo donde la sustancia ósea compacta limita directamente con la cavidad medular. En los mismos lugares por donde la sustancia compacta pasa a la sustancia esponjosa, sus placas comunes internas continúan hacia las placas de las trabéculas de la sustancia esponjosa.
Las placas exteriores están perforadas. canales de perforación (Volkmann), a través del cual los vasos ingresan desde el periostio al hueso. Desde el periostio, las fibras de colágeno penetran en el hueso en diferentes ángulos. Estas fibras se llaman Fibras perforantes (Sharpey). La mayoría de las veces se ramifican solo en la capa exterior de las láminas comunes, pero también pueden penetrar en la capa osteon media, pero nunca entran en las láminas osteon.
En la capa media hay huesos. placas concéntricas (osteónicas) formar osteonas. Entre las osteonas se encuentran placas intermedias (insertar). El grosor y la longitud de las placas óseas varían desde varias decenas hasta cientos de micrómetros. osteonas(Sistemas de Havers) son unidades estructurales de la sustancia compacta del hueso tubular (ver Fig. 8.31, Fig. 8.33). Son formaciones cilíndricas que constan de placas óseas concéntricas, como insertadas entre sí. En las placas óseas y entre ellas se encuentran los cuerpos de las células óseas y sus procesos, incrustados en la sustancia intercelular ósea. Cada osteona está delimitada de las osteonas vecinas por una línea de cementación (fusión) formada por la sustancia fundamental. en el centro

Arroz. 8.33. osteona:
A- microfotografía (tinción según el método Schmorl); b- microscopía electrónica de barrido de un fragmento de hueso (preparación de O. V. Slesarev). 1 - canal de osteonas; 2 - osteocitos (lagunas - b); 3 - placas óseas
El canal de osteona contiene vasos sanguíneos acompañados de tejido conectivo y células osteogénicas.
En la diáfisis de un hueso largo, las osteonas se encuentran predominantemente paralelas al eje longitudinal. Los canales de osteonas se anastomosan entre sí; en los lugares de anastomosis, las placas adyacentes cambian de dirección (v. fig. 8.31). Estos canales se llaman perforador, o nutritivo. Los vasos ubicados en los canales de osteonas se comunican entre sí y con los vasos de la médula ósea y el periostio. La mayor parte de la diáfisis está formada por la sustancia compacta de los huesos tubulares. En la superficie interior del diámetro.
fisis que bordea la cavidad medular, el tejido óseo laminar forma los travesaños óseos del hueso esponjoso. La cavidad de la diáfisis de los huesos tubulares está llena de médula ósea.
Endostio - membrana que cubre el hueso desde el lado de la cavidad medular. En el endostio de la superficie ósea formada, se distingue una línea osmiófila en el borde exterior de la sustancia ósea mineralizada; capa osteoide, que consiste en una sustancia amorfa, fibrillas de colágeno y osteoblastos, capilares sanguíneos y terminaciones nerviosas, una capa de células que separa vagamente el endostio de los elementos de la médula ósea. El espesor del endostio supera las 1-2 micrones, pero es menor que el del periostio.
En áreas de formación ósea activa, el grosor del endostio aumenta de 10 a 20 veces debido a la capa de osteoide debido al aumento de la actividad sintética de los osteoblastos y sus precursores. Durante la remodelación ósea, los osteoclastos se encuentran en el endostio. En el endostio del hueso envejecido, la población de osteoblastos y células progenitoras disminuye, pero aumenta la actividad de los osteoclastos, lo que conduce al adelgazamiento de la capa compacta y a la reestructuración del hueso esponjoso.
Entre el endostio y el periostio existe una cierta microcirculación de líquido y minerales gracias al sistema lacunar-canalicular del tejido óseo.
Vascularización del tejido óseo. Los vasos sanguíneos forman una densa red en la capa interna del periostio. De aquí se originan finas ramas arteriales, que suministran sangre a las osteonas a través de orificios de nutrientes, para luego penetrar en la médula ósea y participar en la formación de la red capilar que la alimenta. Los vasos linfáticos se encuentran principalmente en la capa exterior del periostio.
Inervación del tejido óseo. En el periostio, las fibras nerviosas mielinizadas y amielínicas forman un plexo. Algunas de las fibras acompañan a los vasos sanguíneos y penetran con ellos a través de las aberturas de nutrientes hacia los canales del mismo nombre, luego hacia los canales de osteonas y luego llegan a la médula ósea. Otra parte de las fibras termina en el periostio con ramas nerviosas libres y también participa en la formación de cuerpos encapsulados.
Crecimiento de huesos tubulares. El crecimiento óseo es un proceso muy largo. Comienza en los humanos desde las primeras etapas embrionarias y termina en promedio a la edad de 20 años. Durante todo el período de crecimiento, el hueso aumenta tanto en longitud como en anchura. El crecimiento longitudinal del hueso tubular está garantizado por la presencia de una placa de crecimiento cartilaginosa metaepifisaria, en la que se manifiestan dos procesos histogenéticos opuestos.
Uno es la destrucción de la placa epifisaria y el otro, todo lo contrario, es la reposición constante de tejido cartilaginoso mediante la formación de nuevas células. Sin embargo, con el tiempo, los procesos de destrucción celular comienzan a prevalecer sobre los procesos de neoplasia, como resultado de lo cual la placa cartilaginosa se vuelve más delgada y desaparece. Se detiene el crecimiento óseo en longitud.
En el cartílago metaepifisario se encuentran borde zona, zona células columnares y zona células vesiculares. Zona fronteriza ubicada cerca
epífisis, consta de células redondas y ovaladas y grupos isogénicos únicos que proporcionan conexión entre la placa cartilaginosa y el hueso de la epífisis. En las cavidades entre el hueso y el cartílago hay capilares sanguíneos que nutren las células de las zonas más profundas de la placa cartilaginosa. La zona de células columnares contiene células que proliferan activamente y forman columnas a lo largo del eje longitudinal del hueso y garantizan su crecimiento y longitud. Los extremos proximales de las columnas están formados por células cartilaginosas en proceso de maduración y diferenciación. Son ricos en glucógeno y fosfatasa alcalina. Ambas zonas son más reactivas a la acción de hormonas y otros factores que influyen en los procesos de osificación y crecimiento óseo. La zona de células vesiculares se caracteriza por la hidratación y destrucción de los condrocitos seguida de la osificación endocondral. La parte distal de esta zona limita con la diáfisis, desde donde penetran los capilares sanguíneos y las células osteogénicas. Las columnas de hueso endocondral orientadas longitudinalmente son esencialmente tubos óseos en los que se forman osteonas.
Posteriormente, los centros de osificación de la diáfisis y la epífisis se fusionan y finaliza el crecimiento longitudinal del hueso.
El crecimiento del hueso tubular en anchura se debe al periostio. En el lado del periostio, muy temprano comienzan a formarse capas concéntricas de tejido óseo laminar. Este en aposición el crecimiento continúa hasta que se completa la formación ósea. La cantidad de osteonas inmediatamente después del nacimiento es pequeña, pero a los 25 años su cantidad en los huesos largos de las extremidades aumenta significativamente.
Regeneración. La regeneración fisiológica del tejido óseo se produce lentamente debido a las células osteogénicas del periostio: el endostio y las células osteogénicas en el canal de osteona. La regeneración postraumática del tejido óseo se produce mejor en los casos en que los extremos del hueso roto no están desplazados entre sí. El proceso de osteogénesis está precedido por la formación de callos de tejido conectivo, en cuyo espesor se pueden formar islas cartilaginosas (fig. 8.34). La osificación en este caso se produce según el tipo de osteogénesis secundaria (indirecta). En condiciones de óptima oxigenación tisular, buena reposición y fijación de los extremos del hueso roto, la regeneración se produce sin formación de callos. Sin embargo, antes de que los osteoblastos comiencen a formar hueso, los osteoclastos forman un pequeño espacio entre los extremos acoplados del hueso. Este patrón biológico es la base para el uso por parte de los traumatólogos de dispositivos para el estiramiento gradual de los huesos fusionados durante todo el período de regeneración.
Remodelación ósea y factores que influyen en su estructura.
En el tejido óseo a lo largo de la vida de una persona, se producen procesos interconectados de destrucción y creación, causados por cargas funcionales y otros factores del entorno externo e interno. La reestructuración de las osteonas siempre está asociada con la destrucción de las osteonas primarias y la formación simultánea de nuevas osteonas tanto en el lugar de la destrucción como a partir del periostio. Bajo la influencia de los osteoclastos activados.

Arroz. 8.34. Regeneración postraumática del hueso tubular: A- localización de lesiones; dios- etapas sucesivas de regeneración sin fijación rígida de huesos reducidos (b 1, En 1- fragmentos); d- regeneración tras la fijación de fragmentos. 1 - periostio; 2 - barras transversales de tejido óseo de fibra gruesa; 3 - el tejido conectivo se regenera con islas de tejido cartilaginoso; 4 - el hueso se regenera a partir de tejido óseo fibroso grueso; 5 - línea de fusión (según R.V. Krstic, con modificaciones)
Varios factores, las placas óseas del osteon se destruyen y se forma una cavidad en su lugar. Este proceso se llama reabsorción(del lat. resorción- resorción) del tejido óseo. Los osteoblastos aparecen en la cavidad resultante alrededor del vaso restante y comienza la construcción de nuevas placas, superpuestas concéntricamente entre sí. Así surgen las generaciones secundarias de osteonas. Entre las osteonas se encuentran los restos de osteonas destruidas de generaciones anteriores. El proceso de reestructuración de las osteonas no se detiene incluso después del final del crecimiento óseo.
Entre los factores que influyen en la reestructuración del tejido óseo, juega un papel importante el llamado efecto piezoeléctrico. Resultó que al doblar la placa ósea aparece una cierta diferencia de potencial entre los lados cóncavo y convexo. El primero tiene carga negativa,
y el segundo es positivo. En una superficie cargada negativamente siempre se observa la activación de los osteoblastos y el proceso de formación de hueso nuevo por aposición, mientras que en una superficie cargada positivamente, por el contrario, se observa su reabsorción con la ayuda de los osteoclastos. La creación artificial de una diferencia de potencial conduce al mismo resultado (figura 8.35). El potencial cero, la falta de estrés físico sobre el tejido óseo (inmovilización prolongada, estado de ingravidez, etc.) provocan un aumento de las funciones de los osteoclastos y la excreción de sales.
La estructura del tejido óseo y los huesos está influenciada por las vitaminas (C, A, D), las hormonas tiroideas, paratiroides y otras glándulas endocrinas.
En particular, con una cantidad insuficiente de vitamina C en el cuerpo (por ejemplo, con el escorbuto), se suprime la formación de fibras de colágeno, se debilita la actividad de los osteoblastos y se reduce su actividad fosfatasa, lo que prácticamente conduce al cese de crecimiento óseo debido a la inhibición de la formación de la base orgánica del tejido óseo. En caso de deficiencia de vitamina D (raquitismo), no se produce una calcificación completa de la matriz ósea orgánica, lo que provoca un reblandecimiento de los huesos (osteomalacia). La vitamina A favorece el crecimiento óseo, pero un exceso de esta vitamina aumenta la destrucción del cartílago metaepifisario por los osteoclastos (la zona de crecimiento de los huesos) y ralentiza su alargamiento.
Con un exceso de hormona paratiroidea, paratirina, se observa un aumento de la actividad de los osteoclastos y la resorción ósea. La tirocalcitonina, producida por las células C de la glándula tiroides, actúa de manera diametralmente opuesta, reduciendo la función de los osteoclastos que tienen receptores para esta hormona. Con hipofunción de la glándula tiroides, el crecimiento de los huesos tubulares largos se ralentiza como resultado de la supresión de la actividad de los osteoblastos y la inhibición del proceso de osificación. La regeneración ósea en este caso se produce de forma débil e incompleta. En caso de subdesarrollo testicular o castración prepuberal, se retrasa la osificación de la placa metaepifisaria, por lo que los brazos y las piernas de dicho individuo se vuelven desproporcionadamente largos. Con la falta de estrógeno después de la menopausia, las mujeres a veces desarrollan osteoporosis. Con la pubertad temprana, el crecimiento se detiene debido a la fusión prematura de los huesos diafisarios y epifisarios. Un cierto papel positivo en el crecimiento óseo lo desempeña la hormona somatotrópica de la adenohipófisis, que estimula el desarrollo esquelético proporcional en edades jóvenes (adolescentes) y desproporcionado (acromegalia) en adultos.
Cambios relacionados con la edad. Los tejidos conectivos sufren cambios en estructura, cantidad y composición química con la edad. Con la edad, aumenta la masa total de formaciones de tejido conectivo y el crecimiento del esqueleto óseo. En muchos tipos de estructuras de tejido conectivo, la proporción de tipos de colágeno y glucosaminoglicanos cambia; en particular, contienen más compuestos sulfatados.
Conexiones óseas
Dos huesos pueden tener conexiones. continuo(sindesmosis, sincondrosis y sinostosis) y intermitente(articulaciones).
Conexiones continuas- conexiones con la ayuda de tejido conectivo fibroso denso, cuyos haces en forma de fibras perforantes se introducen en el hueso;

Arroz. 8.35. Efecto piezoeléctrico (explicaciones en el texto): I - diagrama de la organización estructural de las trabéculas óseas; II - activación de osteoclastos y osteoblastos cuando cambia la forma de la trabécula ósea; III - creación artificial de diferencia de potencial (según Yu. I. Afanasyev)
tela nueva. Un ejemplo de tales conexiones son las suturas de los huesos parietales del cráneo, la membrana de tejido conectivo entre los huesos del radio y el cúbito.
Sincondrosis (sínfisis)- conexiones realizadas por cartílago, como los discos intervertebrales. Consisten en un anillo fibroso externo y una parte interna llamada núcleo pulposo. Ambas partes están vagamente separadas y imperceptiblemente se transforman entre sí. El núcleo pulposo se encuentra en la zona interna del disco intervertebral. En diferentes periodos de edad tiene una estructura diferente. A la edad de hasta 2 años, es una cavidad con contenido homogéneo, que contiene solo células individuales. En los años siguientes de vida, esta cavidad se divide en cámaras separadas. A partir de los 6-8 años se nota la aparición y luego aumento del número de fibras de colágeno y células cartilaginosas en el núcleo pulposo. A partir de los 15 años, el crecimiento de fibras y células cartilaginosas se intensifica aún más, y entre los 20 y los 23 años, el núcleo pulposo adquiere el aspecto característico del cartílago fibroso. Un ejemplo de otra conexión más estrecha sería la sínfisis púbica. Las sincondrosis también incluyen conexiones de la epífisis y la diáfisis con la ayuda del cartílago metaepifisario.
sinostosis- articulaciones óseas densas sin tejido conectivo fibroso, como los huesos de la pelvis.
Las articulaciones intermitentes, o articulaciones (diartrosis), consisten en superficies articulares cubiertas de cartílago y, en algunos casos, un menisco intermedio cartilaginoso y una cápsula articular. La cápsula articular consta de una capa fibrosa exterior y una capa sinovial interior. Por este último se entiende una capa de tejido conectivo específicamente diferenciado que contiene vasos sanguíneos y linfáticos, fibras nerviosas y terminaciones. La posición límite de este tejido conectivo, inusual para otros derivados mesenquimales, el constante estiramiento, desplazamiento y presión debido a la participación de la función locomotora de la articulación determina el crecimiento y sus características estructurales.
En la membrana sinovial de mamíferos y humanos, se distinguen dos capas fibrosas elásticas de colágeno (superficial y profunda) y una capa de cobertura que recubre la cavidad (ver Fig. 8.20). No existe un límite definido entre las capas. En las articulaciones grandes existe una capa subsinovial rica en tejido adiposo, que bordea la cápsula fibrosa. Las fibras de colágeno y elásticas de la capa superficial están orientadas en la dirección del eje longitudinal de la articulación. En la capa profunda se encuentran en ángulo con respecto a las fibras de la capa superficial.
La capa que cubre la membrana sinovial está formada por células: los sinoviocitos. Existen sinoviocitos macrófagos y fibroblastos sinoviales, que tienen la capacidad de producir y secretar ácido hialurónico, un componente específico del líquido sinovial.
Los vasos sanguíneos penetran en la membrana sinovial desde los tejidos subyacentes y se distribuyen por todo su espesor, incluida la capa tegumentaria, donde se ubican directamente debajo de los sinoviocitos. Por tanto, la cavidad sinovial está separada del torrente sanguíneo únicamente por las células, la sustancia principal del tejido conectivo y el endotelio de los propios capilares. El endotelio de los hemocapilares de las membranas sinoviales se caracteriza por fenestras y la capacidad de fagocitar. Los capilares linfáticos siempre se encuentran más profundos que los capilares sanguíneos dentro de la capa fibrosa superficial.
La membrana sinovial está ricamente inervada por fibras de naturaleza aferente y eferente (simpática).
Preguntas de control
1. El mesénquima como fuente de desarrollo de los tejidos conectivos: el concepto de diferenciación divergente del mesénquima, características morfológicas del mesénquima.
2. Tejidos conectivos con propiedades especiales: clasificación, topografía del cuerpo, estructura, funciones.
3. Diferencias celulares del tejido conectivo laxo: fuentes de desarrollo, estructura, funciones, participación en la regeneración fisiológica y reparadora.
4. Tejido cartilaginoso: clasificación, topografía, estructura, funciones, regeneración.
5. Tejido óseo: osteogénesis directa e indirecta, estructura, composición celular-diferencial, regeneración.
Histología, embriología, citología: libro de texto / Yu. I. Afanasyev, N. A. Yurina, E. F. Kotovsky, etc. - 6ª ed., revisada. y adicional - 2012. - 800 p. : enfermo.
Lea también...
- Teorías motivacionales. Motivo y motivación. Teorías de la motivación Teorías de la motivación en diversas direcciones psicológicas.
- Propósito de la prueba de ansiedad escolar de Phillips
- Academia Regional Estatal de Samara
- M. V. Koltunova lenguaje y comunicación empresarial. Lenguaje y comunicación empresarial Etiqueta y protocolo de la comunicación empresarial.