Elusaine biosfääri taseme tunnused. Ettekanne teemal “Aine organiseerimise bioloogilise taseme tunnused” Darwini evolutsiooniteooria
Naturalistlik bioloogia Aristoteles: -Jagas loomariigi kahte rühma: verega ja vereta. - Inimene on vereloomade peal (antropotsentrism). K. Linnaeus: -töötas välja kõikide loomade ja taimede harmoonilise hierarhia (liik - perekond - järg - klass), -võtis kasutusele täpse terminoloogia taimede ja loomade kirjeldamiseks.


Evolutsioonibioloogia Küsimus elu tekkest ja olemusest. J. B. Lamarck pakkus välja esimese evolutsiooniteooria aastal 1809. J. Cuvier pakkus välja katastroofide teooria. Charles Darwini evolutsiooniteooria 1859. aastal evolutsiooniteooria 1859. aastal Kaasaegne (sünteetiline) evolutsiooniteooria (esindab geneetika ja darvinismi sünteesi).



Molekulaargeneetiline tase Organismide eluprotsesside aluseks olevate biopolümeeride (valgud, nukleiinhapped, polüsahhariidid) jne funktsioneerimise tase. Elementaarne struktuuriüksus on geen Päriliku informatsiooni kandjaks on DNA molekul.





Nukleiinhapped Komplekssed orgaanilised ühendid, mis on fosforit sisaldavad biopolümeerid (polünukleotiidid). Tüübid: desoksüribonukleiinhape (DNA) ja ribonukleiinhape (RNA). Organismi geneetiline informatsioon salvestatakse DNA molekulidesse. Neil on molekulaarse dissümmeetria (asümmeetria) ehk molekulaarse kiraalsuse omadus – nad on optiliselt aktiivsed.

DNA koosneb kahest ahelast, mis on keerdunud topeltheeliksiks. RNA sisaldab 4-6 tuhat üksikut nukleotiidi, DNA - tuhandeid. Geen on DNA või RNA molekuli osa.

Rakutasand Sellel tasandil toimub elutähtsate protsesside ruumiline piiritlemine ja järjestamine tänu funktsioonide jagunemisele konkreetsete struktuuride vahel. Kõigi elusorganismide põhiline struktuurne ja funktsionaalne üksus on rakk. Sellest organiseerituse tasemest sai alguse meie planeedi elulugu.



Kõik elusorganismid koosnevad rakkudest ja nende ainevahetusproduktidest. Uued rakud moodustuvad juba olemasolevate rakkude jagunemisel. Kõik rakud on keemilise koostise ja ainevahetuse poolest sarnased. Organismi kui terviku tegevus koosneb üksikute rakkude tegevusest ja vastasmõjust.

1830. aastatel. Rakutuum avastati ja kirjeldati. Kõik rakud koosnevad: 1) plasmamembraanist, mis kontrollib ainete üleminekut keskkond puuri ja tagasi; 2) mitmekesise ehitusega tsütoplasmad; 3) rakutuum, mis sisaldab geneetilist informatsiooni.




Ontogeneetiline (organismi) tasand Organism on terviklik ainurakne või mitmerakuline elusüsteem, mis on võimeline iseseisvalt eksisteerima. Ontogenees on organismi individuaalse arengu protsess sünnist surmani, päriliku teabe realiseerimise protsess.





Populatsioon on ühte liiki isendite kogum, mis hõivab teatud territooriumi, taastoodab end pika aja jooksul ja millel on ühine geneetiline reserv. Liik on struktuurilt ja füsioloogilistelt omadustelt sarnaste isendite kogum, millel on ühine päritolu ja mis võivad vabalt ristuda ja anda viljakaid järglasi.



Biogeotsenootiline tasand Biogeotsenoos ehk ökoloogiline süsteem (ökosüsteem) on aine-, energia- ja infovahetuse kaudu omavahel seotud biootiliste ja abiootiliste elementide kogum, mille raames saab toimuda ainete ringlemine looduses.

Biogeocenoos on terviklik isereguleeruv süsteem, mis koosneb: 1) tootjatest (tootjatest), kes töötlevad vahetult elutut ainet (vetikad, taimed, mikroorganismid); 2) esimest järku tarbijad - ainet ja energiat saadakse tootjate (rohutoiduliste) kasutamise kaudu; 3) teist järku tarbijad (kiskjad jne); 4) koristajad (saprofüüdid ja saprofaagid), toituvad surnud loomadest; 5) lagundajad on rühm baktereid ja seeni, mis lagundavad orgaanilise aine jääke.




"Biosfäär ja tsivilisatsioon" – abiootilised tegurid. Ökoloogia põhimõisted. Keskkonnategur. Taimtoidulised. Ameerika teadlane. Raamat V.I. Vernadski "Biosfäär". Inimtegevus. Kasvuhooneefekt. Ökoloogiline nišš. Piiravad tegurid. Biosfääri alumine piir. Liigne vesi. Eduard Suess. Autotroofid. Antropogeenne tegur. Veetarbimine. Rahvastiku kasv. Vaate asend ruumis. Kompenseerivad omadused.
"Biosfääri mõiste" - Inimese reaktsioonid muutustele biosfääris. Malaaria. Biosfääri evolutsioon. Elusaine biosfääris. Filmid elust ookeanis. Jean-Baptiste Lamarcki portree. Sargassumi vetikad. Kuidas filosoofid noosfääri esindavad. Orgaaniliste ja anorgaaniliste ainete lagunemine. Näide inimese ebaõnnestunud sekkumisest. Noosfäär. Elusorganismid. Spetsiaalne keemiline koostis. Lämmastiku tsükkel. Biosfääri koostis. Riftii. Anaeroobsed bakterid.
“Biosfäär kui globaalne ökosüsteem” – Biosfäär kui globaalne biosüsteem ja ökosüsteem. Elutu loodus. Organismide elukeskkond Maal. Inimene kui biosfääri elanik. Maa kest. Bioloogiline tsükkel. Keskkonnategurid. Elusorganismid. Inimene. Biosfäär kui globaalne biosüsteem. Elusaine biosfääri taseme tunnused.
"Biosfäär on Maa elav kest" - elutu loodus. Meie planeedi iidsete elanike välimus. Elusorganismid. Kivid. Taimkate. Soe. Biosfäär. Maa. Rohelised taimed. Olendid.
"Biosfääri koostis ja struktuur" - Biosfääri piirid. Evolutsiooniline seisund. Vernadski. Piirav tegur. Hüdrosfäär. Maa kest. Elav aine. Litosfäär. Osoonikiht. Noosfäär. Biosfääri struktuur. Biosfäär. Atmosfäär.
"Biosfääri uurimine" - bakterid, eosed ja õietolm. Interaktsioon. Elu päritolu Maal. Mis on ligikaudu planeet Maa vanus. Elujõulisus. Kõik organismid on ühendatud 4 eluslooduse kuningriiki. Organismide mitmekesisus. 40 tuhat aastat tagasi ilmus moodne inimene. Mitu liiki seeni on? Biosfääri piirid. Kontrolli ennast. Millega varustab biosfäär hüdrosfääri? Mäng "Biosfäär". Organismide mitmekesisus Maal.
Föderaalne tervishoiu- ja sotsiaalamet
Test bioloogias
Elusaine kvalitatiivsed omadused. Elusolendite organiseerituse tasemed.
Raku keemiline koostis (valgud, nende struktuur ja funktsioonid)
Lõpetanud õpilane
1. aasta 195 rühm
kirjavahetusosakond
Farmaatsiateaduskond
Tšeljabinsk 2009
Elusaine kvalitatiivsed omadused. Elusolendite organiseerituse tasemed
Iga elussüsteem, olenemata sellest, kui keeruline see on, koosneb bioloogilistest makromolekulidest: nukleiinhapetest, valkudest, polüsahhariididest, aga ka muudest olulistest orgaanilistest ainetest. Sellelt tasandilt algavad organismi erinevad elutähtsad protsessid: ainevahetus ja energia muundamine, päriliku info edastamine jne.
Mitmerakuliste organismide rakud moodustavad kudesid – struktuurilt ja funktsioonilt sarnaseid rakusüsteeme ning nendega seotud rakkudevahelisi aineid. Kuded integreeritakse suuremateks funktsionaalseteks üksusteks, mida nimetatakse organiteks. Siseorganid on loomadele iseloomulikud; siin on nad osa organsüsteemidest (hingamisteede, närvisüsteemi jne). Näiteks seedesüsteem: suuõõs, neel, söögitoru, magu, kaksteistsõrmiksool, peensool, käärsool, pärak. Selline spetsialiseerumine ühelt poolt parandab organismi kui terviku talitlust, teisalt aga nõuab erinevate kudede ja elundite suuremat koordinatsiooni ja integratsiooni.
Rakk on struktuurne ja funktsionaalne üksus, samuti kõigi Maal elavate elusorganismide arenguüksus. Rakutasandil on info edastamine ning ainete ja energia muundamine seotud.
Organismi tasandi elementaarüksus on indiviid, keda peetakse arengus – tekkehetkest eksistentsi lõpuni – elavaks süsteemiks. Tekivad elundisüsteemid, mis on spetsialiseerunud erinevate funktsioonide täitmisele.
Sama liigi organismide kogum, mida ühendab ühine elupaik, milles luuakse populatsioon - organismiülene süsteem. Selles süsteemis viiakse läbi elementaarsed evolutsioonilised teisendused.
Biogeocenoos – organismide kogum erinevad tüübid ja organisatsiooni erinev keerukus koos keskkonnateguritega. Erinevate süstemaatiliste rühmade organismide ühise ajaloolise arengu käigus moodustuvad dünaamilised, stabiilsed kooslused.
Biosfäär on kõigi biogeotsenooside kogum, süsteem, mis hõlmab kõiki meie planeedi elunähtusi. Sellel tasemel toimub ainete ringlus ja energia muundumine, mis on seotud kõigi elusorganismide elutegevusega.
Tabel 1. Elusaine organiseerituse tasemed
Molekulaarne
Elusolendite organiseerituse esialgne tase. Uurimisobjektiks on nukleiinhapete, valkude, süsivesikute, lipiidide ja teiste bioloogiliste molekulide molekulid, s.o. rakus leiduvad molekulid. Iga elussüsteem, olenemata sellest, kui keeruline see on, koosneb bioloogilistest makromolekulidest: nukleiinhapetest, valkudest, polüsahhariididest, aga ka muudest olulistest orgaanilistest ainetest. Sellelt tasandilt algavad organismi erinevad elutähtsad protsessid: ainevahetus ja energia muundamine, päriliku info edastamine jne.
Mobiilne
Sõltumatute organismidena toimivate rakkude (bakterid, algloomad ja mõned muud organismid) ning mitmerakulisi organisme moodustavate rakkude uurimine.
Kangas
Ühise päritoluga ja sarnaseid funktsioone täitvad rakud moodustavad kudesid. Erinevate omadustega loomseid ja taimseid kudesid on mitut tüüpi.
Organ
Organismides, alustades koelenteraatidest, moodustuvad organid (organisüsteemid), sageli erinevat tüüpi kudedest.
Organism
Seda taset esindavad ühe- ja mitmerakulised organismid.
Populatsioon-liigid
Teatud piirkondades koos elavad sama liigi organismid moodustavad populatsiooni. Nüüd on Maal umbes 500 tuhat taimeliiki ja umbes 1,5 miljonit loomaliiki.
Biogeotsenootiline
Seda esindab erinevate liikide organismide kogum, mis sõltuvad üksteisest ühel või teisel määral.
Biosfäär
Elusolendite organiseerimise kõrgeim vorm. Hõlmab kõiki üldise ainevahetuse ja energia muundamisega seotud biogeotsenoose.
Kõik need tasemed on üsna spetsiifilised, neil on oma mustrid, oma uurimismeetodid. Võib isegi välja tuua teadused, mis viivad oma uurimistööd läbi elusolendite teatud organiseerituse tasemel. Näiteks molekulaarsel tasandil uurivad elusolendeid sellised teadused nagu molekulaarbioloogia, bioorgaaniline keemia, bioloogiline termodünaamika, molekulaargeneetika jne. Kuigi elusolendite organiseerituse tasandid eristuvad, on need omavahel tihedalt seotud ja voolavad üksteisest, mis räägib eluslooduse terviklikkusest.
Rakumembraan. Lahtri pinnaaparaat, selle põhiosad, otstarve
Elav rakk on põhiosake elusaine struktuurid. See on kõige lihtsam süsteem, millel on kõik elusolendite omadused, sealhulgas võime edastada geneetilist teavet. Rakuteooria lõid Saksa teadlased Theodor Schwann ja Matthias Schleiden. Selle peamine seisukoht on väide, et kõik taime- ja loomaorganismid koosnevad sarnase struktuuriga rakkudest. Tsütoloogiaalased uuringud on näidanud, et kõik rakud teostavad ainevahetust, on võimelised isereguleeruma ja edastama pärilikku teavet. Iga raku elutsükkel lõpeb kas jagunemisega ja elu jätkamisega uuenenud kujul või surmaga. Samal ajal selgus, et rakud on väga mitmekesised, nad võivad eksisteerida üherakuliste organismidena või osana mitmerakulistest. Rakkude eluiga ei tohi ületada mitut päeva või võib langeda kokku organismi elueaga. Rakkude suurused on väga erinevad: 0,001–10 cm Rakud moodustavad kudesid, kehasüsteemideks nimetatakse mitut tüüpi kudesid – organeid, elundirühmi, mis on seotud mõne levinud probleemi lahendamisega. Rakkudel on keeruline struktuur. See on väliskeskkonnast eraldatud kestaga, mis, olles lahti ja lahti, tagab raku vastasmõju välismaailmaga, aine-, energia- ja infovahetuse sellega. Rakkude metabolism on aluseks veel ühele nende kõige olulisemale omadusele - raku sisekeskkonna tingimuste stabiilsuse ja stabiilsuse säilitamisele. Seda rakkude omadust, mis on omane kogu elussüsteemile, nimetatakse homöostaasiks. Homöostaasi, see tähendab raku koostise püsivust, säilitab ainevahetus, see tähendab ainevahetus. Ainevahetus on keeruline, mitmeetapiline protsess, mis hõlmab tooraine viimist rakku, nendest energia ja valkude tootmist ning toodetud kasulike saaduste, energia ja jäätmete viimist rakust keskkonda.
Rakumembraan on rakumembraan, mis täidab järgmisi funktsioone:
raku sisu ja väliskeskkonna eraldamine;
raku ja keskkonna vahelise ainevahetuse reguleerimine;
mõnede biokeemiliste reaktsioonide koht (sealhulgas fotosüntees, oksüdatiivne fosforüülimine);
rakkude ühendamine kudedeks.
Membraanid jagunevad plasmateks (rakumembraanid) ja välisteks. Plasmamembraani kõige olulisem omadus on poolläbilaskvus, see tähendab võime lasta läbi ainult teatud aineid. Glükoos, aminohapped, rasvhapped ja ioonid difundeeruvad selle kaudu aeglaselt ning membraanid ise saavad difusiooniprotsessi aktiivselt reguleerida.
Kaasaegsete andmete kohaselt on plasmamembraanid lipoproteiinide struktuurid. Lipiidid moodustavad spontaanselt kaksikkihi ja membraanivalgud "hõljuvad" selles. Membraanid sisaldavad mitut tuhat erinevat valku: struktuurseid, transportereid, ensüüme jt. Eeldatakse, et valgumolekulide vahel on poorid, millest hüdrofiilsed ained läbi pääsevad (lipiidne kaksikkiht takistab nende otsest tungimist rakku). Glükosüülrühmad on kinnitunud membraani pinnale mõnele molekulile, mis osalevad koe moodustumise käigus rakkude äratundmise protsessis.
Erinevad tüübid membraanid erinevad oma paksuse poolest (tavaliselt jääb see vahemikku 5–10 nm). Lipiidide kaksikkihi konsistents meenutab oliiviõli. Sõltuvalt välistingimustest (regulaatoriks on kolesterool) võib kaksikkihi struktuur muutuda nii, et see muutub vedelamaks (sellest sõltub membraani aktiivsus).
Oluline teema on ainete transport läbi plasmamembraanide. See on vajalik toitainete rakku toimetamiseks, mürgiste jäätmete eemaldamiseks ning gradientide tekitamiseks närvi- ja lihastegevuse säilitamiseks. Ainete transportimiseks läbi membraani on olemas järgmised mehhanismid:
difusioon (gaasid, rasvlahustuvad molekulid tungivad otse läbi plasmamembraani); hõlbustatud difusiooniga läbib vees lahustuv aine membraani läbi konkreetse molekuli loodud spetsiaalse kanali;
osmoos (vee difusioon läbi poolläbilaskvate membraanide);
aktiivne transport (molekulide ülekandmine madalama kontsentratsiooniga piirkonnast kõrgema kontsentratsiooniga piirkonda, näiteks spetsiaalsete transpordivalkude kaudu, nõuab ATP energiat);
Endotsütoosi ajal moodustab membraan invaginatsioone, mis seejärel muundatakse vesiikuliteks või vakuoolideks. On fagotsütoos - tahkete osakeste imendumine (näiteks vere leukotsüütide poolt) - ja pinotsütoos - vedelike imendumine;
eksotsütoos on endotsütoosi pöördprotsess; Rakkudest eemaldatakse seedimata tahkete osakeste jäänused ja vedelad eritised.
Ülemembraansed struktuurid võivad paikneda raku plasmamembraani kohal. Nende struktuur on märja klassifikatsiooni tunnus. Loomadel on see glükokalüks (valgu-süsivesikute kompleks), taimedes, seentes ja bakterites on see rakusein. Taimede rakuseinas on tselluloos, seened - kitiin, bakterid - valgu-polüsahhariidi kompleks mureiin.
Rakupinna aparaadi (SAC) aluseks on välimine rakumembraan ehk plasmalemma. Lisaks plasmamembraanile on PAA-l membraaniülene kompleks ja eukarüootides ka submembraanne kompleks.
Plasmalemma (kreeka keelest plasma - moodustumine ja lemma - kest, koorik) peamised biokeemilised komponendid on lipiidid ja valgud. Nende kvantitatiivne suhe enamikus eukarüootides on 1:1 ja prokarüootides on plasmalemmas ülekaalus valgud. Väike kogus süsivesikuid leidub raku välismembraanis ja rasvataolisi ühendeid (imetajatel - kolesterool, rasvlahustuvad vitamiinid).
Rakupinna aparaadi supramembraanset kompleksi iseloomustavad mitmesugused struktuurid. Prokarüootides esindab supramembraanset kompleksi enamikul juhtudel erineva paksusega rakusein, mille aluseks on kompleksne glükoproteiin mureiin (arhebakterites - pseudomureiin). Paljude eubakterite puhul koosneb supramembraanse kompleksi välimine osa teisest suure lipopolüsahhariidide sisaldusega membraanist. Eukarüootides on supramembraanse kompleksi universaalseks komponendiks süsivesikud - plasmalemma glükolipiidide ja glükoproteiinide komponendid. Selle tõttu hakati teda algselt nimetama glükokalüksiks (kreeka keelest glycos – magus, süsivesikuline ja lat. callum – paks nahk, kest). Lisaks süsivesikutele sisaldab glükokalüks perifeerseid valke, mis asuvad bilipiidkihi kohal. Supramembraanse kompleksi keerukamaid variante leidub taimedes (tselluloosist rakusein), seentes ja lülijalgsetes (kitiinist väliskate).
Submembraani (ladina keelest sub-under) kompleks on iseloomulik ainult eukarüootsetele rakkudele. See koosneb mitmesugustest proteiinniiditaolistest struktuuridest: õhukesed fibrillid (ladina fibrill - kiud, niit), mikrofibrillid (kreeka keelest micros - väikesed), skeleti (kreeka skeleti - kuivatatud) fibrillid ja mikrotuubulid. Need on omavahel seotud valkude kaudu ja moodustavad raku luu-lihaskonna aparatuuri. Submembraanne kompleks interakteerub plasmalemma valkudega, mis omakorda on seotud supramembraanse kompleksiga. Selle tulemusena on PAK struktuurselt terviklik süsteem. See võimaldab tal täita raku jaoks olulisi funktsioone: isoleerimine, transport, katalüütiline, retseptori signaalimine ja kontakt.
Raku keemiline koostis (valgud, nende struktuur ja funktsioonid)
Rakus toimuvad keemilised protsessid on selle elu, arengu ja toimimise üks peamisi tingimusi.
PAGE_BREAK--
Kõik taime- ja loomaorganismide rakud, aga ka mikroorganismid on keemilise koostise poolest sarnased, mis näitab ühtsust. orgaaniline maailm.
Mendelejevi perioodilisuse tabeli 109 elemendist leiti rakkudest märkimisväärne enamus. Mõned elemendid sisalduvad rakkudes suhteliselt suurtes kogustes, teised väikestes kogustes (tabel 2).
Tabel 2. Sisukord keemilised elemendid puuris
Elemendid
Kogus (%)
Elemendid
Kogus (%)
Hapnik
Raku ainete hulgas on esikohal vesi. See moodustab peaaegu 80% raku massist. Vesi on raku kõige olulisem komponent, mitte ainult koguseliselt. See mängib raku elus olulist ja mitmekesist rolli.
Vesi määrab füüsikalised omadused rakud - selle maht, elastsus. Vesi omab suurt tähtsust orgaaniliste ainete molekulide struktuuri, eriti valkude struktuuri kujunemisel, mis on vajalik nende funktsioonide täitmiseks. Vee tähtsus lahustina on suur: väliskeskkonnast satuvad rakku paljud ained vesilahuses, vesilahuses eemaldatakse rakust jääkained. Lõpuks on vesi paljudes keemilistes reaktsioonides (valkude, süsivesikute, rasvade jne lagunemine) otsene osaline.
Vee bioloogilise rolli määravad selle molekulaarstruktuuri iseärasused ja molekulide polaarsus.
Raku anorgaaniliste ainete hulka kuuluvad lisaks veele ka soolad. Elutähtsate protsesside jaoks on soolades sisalduvateks olulisemateks katioonideks K+, Na+, Ca2+, Mg2+ ja olulisemateks anioonideks HPO4-, H2PO4-, Cl-, HCO3-.
Katioonide ja anioonide kontsentratsioon rakus ja selle elupaigas on reeglina järsult erinev. Kuni rakk on elus, säilib ioonide suhe rakus ja väljaspool seda kindlalt. Pärast rakusurma ioonide sisaldus rakus ja keskkonnas ühtlustub kiiresti. Rakus sisalduvatel ioonidel on suur tähtsus nii raku normaalseks funktsioneerimiseks kui ka rakusisese pideva reaktsiooni hoidmiseks. Hoolimata asjaolust, et eluprotsessis tekivad pidevalt happed ja leelised, on raku normaalne reaktsioon kergelt aluseline, peaaegu neutraalne.
Anorgaanilised ained sisalduvad rakus mitte ainult lahustunud, vaid ka tahkes olekus. Eelkõige annab luukoe tugevuse ja kõvaduse kaltsiumfosfaat ning molluskite kestadele kaltsiumkarbonaat.
Orgaanilised ained moodustavad umbes 20-30% raku koostisest.
Biopolümeeride hulka kuuluvad süsivesikud ja valgud. Süsivesikud sisaldavad süsiniku, hapniku ja vesiniku aatomeid. Seal on lihtsad ja komplekssed süsivesikud. Lihtne – monosahhariidid. Kompleks – polümeerid, mille monomeerideks on monosahhariidid (oligosahhariidid ja polüsahhariidid). Monomeeriühikute arvu suurenedes polüsahhariidide lahustuvus väheneb ja magus maitse kaob.
Monosahhariidid on tahked, värvitud kristalsed ained, mis lahustuvad vees hästi ja orgaanilistes lahustites väga halvasti (või üldse mitte). Monosahhariidide hulka kuuluvad trioosid, tetroosid, pentoosid ja heksoosid. Oligosahhariididest on levinumad disahhariidid (maltoos, laktoos, sahharoos). Polüsahhariide leidub kõige sagedamini looduses (tselluloos, tärklis, kitiin, glükogeen). Nende monomeerid on glükoosi molekulid. Nad lahustuvad osaliselt vees, paisudes, moodustades kolloidseid lahuseid.
Lipiidid on vees lahustumatud rasvad ja rasvataolised ained, mis koosnevad glütseroolist ja suure molekulmassiga rasvhapped. Rasvad on kolmehüdroksüülse alkoholi glütserooli ja kõrgemate rasvhapete estrid. Loomseid rasvu leidub piimas, lihas ja nahaaluskoes. Taimedes - seemnetes ja viljades. Lisaks rasvadele sisaldavad rakud ka nende derivaate – steroide (kolesterool, hormoonid ja rasvlahustuvad vitamiinid A, D, K, E, F).
Lipiidid on:
rakumembraanide ja raku organellide struktuurielemendid;
energiamaterjal (1g rasva oksüdeerumisel vabaneb 39 kJ energiat);
varuained;
täitma kaitsefunktsiooni (mere- ja polaarloomadel);
mõjutada närvisüsteemi toimimist;
veeallikas organismile (1 kg oksüdeerituna annab 1,1 kg vett).
Nukleiinhapped. Nimetus “nukleiinhapped” tuleb ladinakeelsest sõnast “tuum”, s.o. tuum: need avastati esmakordselt raku tuumades. Nukleiinhapete bioloogiline tähtsus on väga suur. Neil on keskne roll raku pärilike omaduste säilitamisel ja edasikandmisel, mistõttu neid sageli nimetatakse pärilikkuse aineteks. Nukleiinhapped tagavad valkude sünteesi rakus täpselt samamoodi nagu emarakus ja päriliku info edastamise. Nukleiinhappeid on kahte tüüpi – desoksüribonukleiinhape (DNA) ja ribonukleiinhape (RNA).
DNA molekul koosneb kahest spiraalselt keerdunud ahelast. DNA on polümeer, mille monomeerideks on nukleotiidid. Nukleotiidid on ühendid, mis koosnevad fosforhappe molekulist, süsivesikute desoksüriboosist ja lämmastiku alusest. DNA-l on nelja tüüpi lämmastiku aluseid: adeniin (A), guaniin (G), tsütosiin (C), tümiin (T). Iga DNA ahel on polünukleotiid, mis koosneb mitmekümnest tuhandest nukleotiidist. DNA kahekordistumine – reduplikatsioon – tagab päriliku informatsiooni ülekande emarakust tütarrakkudesse.
RNA on polümeer, mis on struktuurilt sarnane ühe DNA ahelaga, kuid väiksema suurusega. RNA monomeerid on nukleotiidid, mis koosnevad fosforhappest, süsivesikute riboosist ja lämmastiku alusest. Tümiini asemel sisaldab RNA uratsiili. Tuntud on kolme tüüpi RNA-d: messenger RNA (i-RNA) – edastab DNA molekulilt infot valgu struktuuri kohta; transport (t-RNA) - transpordib aminohappeid valgusünteesi kohta; ribosomaalne (r-RNA) – leidub ribosoomides, osaleb ribosoomi struktuuri säilitamises.
Väga oluline roll Raku bioenergeetikas mängib rolli adenüülnukleotiid, millele on kinnitunud kaks fosforhappejääki. Seda ainet nimetatakse adenosiintrifosforhappeks (ATP). ATP on universaalne bioloogiline energiaakumulaator: päikese valgusenergia ja tarbitavas toidus sisalduv energia salvestub ATP molekulidesse. ATP on ebastabiilne struktuur, kui ATP muundub ADP-ks (adenosiindifosfaadiks), vabaneb 40 kJ energiat. ATP-d toodetakse loomarakkude mitokondrites ja fotosünteesi käigus taimede kloroplastides. ATP energiaga tehakse keemilist (valkude, rasvade, süsivesikute, nukleiinhapete süntees), mehaanilist (liikumine, lihastöö) tööd, muundatakse elektri- või valgusenergiaks (elektriraiade, angerjate, putukate kuma) energiaks.
Valgud on mitteperioodilised polümeerid, mille monomeerideks on aminohapped. Kõik valgud sisaldavad süsiniku, vesiniku, hapniku ja lämmastiku aatomeid. Paljud valgud sisaldavad ka väävliaatomeid. On valke, mis sisaldavad ka metalliaatomeid – raud, tsink, vask. Happeliste ja aluseliste rühmade olemasolu määrab aminohapete kõrge reaktsioonivõime. Ühe aminohappe aminorühmast ja teise karboksüülrühmast vabaneb veemolekul ja vabanenud elektronid moodustavad peptiidsideme: CO-NN (selle avastas 1888. aastal professor A. Ya. Danilevsky), mistõttu valke nimetatakse polüpeptiidideks. Valgu molekulid on makromolekulid. Tuntud on palju aminohappeid. Kuid ainult 20 aminohapet on tuntud mis tahes looduslike valkude – loomsete, taimsete, mikroobsete, viiruslike – monomeeridena. Neid kutsuti "maagiaks". Asjaolu, et kõigi organismide valgud on üles ehitatud samadest aminohapetest, on järjekordne tõend Maa elusmaailma ühtsusest.
Valgumolekulide struktuuris on 4 organiseerituse taset:
1. Primaarstruktuur - aminohapete polüpeptiidahel, mis on kovalentsete peptiidsidemetega seotud kindlas järjestuses.
2. Sekundaarne struktuur - polüpeptiid ahel heeliksi kujul. Külgnevate pöörete peptiidsidemete ja teiste aatomite vahel tekib arvukalt vesiniksidemeid, mis annavad tugeva struktuuri.
3. Tertsiaarne struktuur – igale valgule omane konfiguratsioon – gloobul. Seda hoiavad madala tugevusega hüdrofoobsed sidemed või adhesioonijõud mittepolaarsete radikaalide vahel, mida leidub paljudes aminohapetes. Samuti on olemas kovalentsed S-S sidemed, mis tekivad väävlit sisaldava aminohappe tsüsteiini kaugel asuvate radikaalide vahel.
4. Kvaternaarne struktuur tekib siis, kui mitmed makromolekulid ühinevad agregaatideks. Seega on hemoglobiin inimese veres nelja makromolekuli agregaat.
Valgu loomuliku struktuuri rikkumist nimetatakse denaturatsiooniks. See tekib kõrge temperatuuri, kemikaalide, kiirgusenergia ja muude tegurite mõjul.
Valkude roll rakkude ja organismide elus:
ehitus (struktuurne) - valgud - keha ehitusmaterjal (kestad, membraanid, organellid, koed, elundid);
katalüütiline funktsioon – ensüümid, mis kiirendavad reaktsioone sadu miljoneid kordi;
lihas-skeleti funktsioon - valgud, mis moodustavad skeleti luud ja kõõlused; lippude, ripsmete liikumine, lihaste kontraktsioon;
transpordifunktsioon - vere hemoglobiin;
kaitsev - vere antikehad neutraliseerivad võõrkehi;
energiafunktsioon - valgu lõhustamisel vabaneb 1 g 17,6 kJ energiat;
regulatiivne ja hormonaalne - valgud on osa paljudest hormoonidest ja osalevad keha eluprotsesside reguleerimises;
retseptor – valgud viivad läbi üksikute ainete selektiivse äratundmise ja molekulidega seondumise protsessi.
Ainevahetus rakus. Fotosüntees. Kemosüntees
Iga organismi olemasolu eelduseks on pidev toitainete vool ja rakkudes toimuvate keemiliste reaktsioonide lõpp-produktide pidev vabanemine. Toitaineid kasutavad organismid keemiliste elementide (peamiselt süsinikuaatomite) aatomite allikana, millest ehitatakse või uuendatakse kõiki struktuure. Lisaks toitainetele saab organism ka vett, hapnikku ja mineraalsooli.
Rakkudesse sisenevad (või fotosünteesi käigus sünteesitud) orgaanilised ained lagundatakse ehitusplokkideks – monomeerideks ja saadetakse kõikidesse keharakkudesse. Osa nende ainete molekule kulub konkreetsele organismile omaste spetsiifiliste orgaaniliste ainete sünteesiks. Rakud sünteesivad valke, lipiide, süsivesikuid, nukleiinhappeid ja muid aineid, mis täidavad erinevaid funktsioone (ehituslik, katalüütiline, reguleeriv, kaitsev jne).
Teine osa rakkudesse sattuvatest madalmolekulaarsetest orgaanilistest ühenditest läheb ATP moodustamiseks, mille molekulid sisaldavad otseselt töö tegemiseks mõeldud energiat. Energia on vajalik kõigi keha spetsiifiliste ainete sünteesiks, selle kõrgelt korrastatud organiseerituse säilitamiseks, ainete aktiivseks transpordiks rakkude sees, ühest rakust teise, ühest kehaosast teise, närviimpulsside edastamiseks, organismide liikumiseks, püsiva kehatemperatuuri hoidmiseks (lindudel ja imetajatel) ja muudel eesmärkidel.
Ainete muundumisel rakkudes tekivad ainevahetuse lõpp-produktid, mis võivad olla organismile mürgised ja eemaldatakse organismist (näiteks ammoniaak). Seega tarbivad kõik elusorganismid pidevalt keskkonnast teatud aineid, muundavad neid ja lasevad keskkonda lõppprodukte.
Jätkamine
--PAGE_BREAK--
Kehas toimuvate keemiliste reaktsioonide kogumit nimetatakse ainevahetuseks või ainevahetuseks. Sõltuvalt protsesside üldisest suunast eristatakse katabolismi ja anabolismi.
Katabolism (dissimilatsioon) on reaktsioonide kogum, mis viib lihtsate ühendite moodustumiseni keerukamatest ühenditest. Kataboolsed reaktsioonid hõlmavad näiteks polümeeride hüdrolüüsi reaktsioone monomeerideks ja viimaste lagunemist süsinikdioksiidiks, veeks, ammoniaagiks, s.o. energia metabolismi reaktsioonid, mille käigus toimub orgaaniliste ainete oksüdatsioon ja ATP süntees.
Anabolism (assimilatsioon) on reaktsioonide kogum keerukate orgaaniliste ainete sünteesimiseks lihtsamatest. Siia kuuluvad näiteks lämmastiku sidumine ja valkude biosüntees, süsivesikute süntees süsihappegaasist ja veest fotosünteesi käigus, polüsahhariidide, lipiidide, nukleotiidide, DNA, RNA ja muude ainete süntees.
Ainete sünteesi elusorganismide rakkudes nimetatakse sageli plastiliseks ainevahetuseks ning ainete lagunemist ja nende oksüdatsiooni, millega kaasneb ATP süntees, energia metabolismiks. Mõlemad ainevahetuse tüübid moodustavad mistahes raku ja seega iga organismi elutegevuse aluse ning on üksteisega tihedalt seotud. Ühest küljest nõuavad kõik plastilised vahetusreaktsioonid energiakulu. Teisest küljest on energia metabolismi reaktsioonide läbiviimiseks vajalik ensüümide pidev süntees, kuna nende eluiga on lühike. Lisaks tekivad plastilise ainevahetuse käigus (näiteks fotosünteesi käigus) hingamiseks kasutatavad ained.
Fotosüntees on orgaanilise aine moodustumine süsinikdioksiidist ja veest valguse käes fotosünteetiliste pigmentide (taimedes klorofüll, bakterites bakterioklorofüll ja bakteriorodopsiin) osalusel. Kaasaegses taimefüsioloogias mõistetakse fotosünteesi all sagedamini fotoautotroofset funktsiooni - valguskvantide energia neeldumise, muundamise ja kasutamise protsesside kogumit erinevates endergoonilistes reaktsioonides, sealhulgas süsinikdioksiidi muundamisel orgaanilisteks aineteks.
Fotosüntees on peamine bioloogilise energia allikas. Fotosünteesivad autotroofid kasutavad seda orgaaniliste ainete sünteesimiseks keemiliste sidemete kujul, vabastades seda hingamis- ja fermentatsiooniprotsessides. Ka inimkonna fossiilkütuste (kivisüsi, nafta, maagaas, turvas) põletamisel saadud energia salvestub fotosünteesi protsessis.
Fotosüntees on anorgaanilise süsiniku peamine sisend bioloogilisse tsüklisse. Kogu atmosfääris leiduv vaba hapnik on biogeenset päritolu ja on fotosünteesi kõrvalsaadus. Oksüdeeriva atmosfääri tekkimine (hapnikukatastroof) muutis täielikult maapinna seisukorda, muutis võimalikuks hingamise ilmnemise ja võimaldas hiljem, pärast osoonikihi tekkimist, elul maale jõuda.
Kemosüntees on autotroofse toitumise meetod, mille puhul CO2-st orgaaniliste ainete sünteesi energiaallikaks on anorgaaniliste ühendite oksüdatsioonireaktsioonid. Seda tüüpi energiatootmist kasutavad ainult bakterid. Kemosünteesi fenomeni avastas 1887. aastal vene teadlane S.N. Vinogradski.
Tuleb märkida, et anorgaaniliste ühendite oksüdatsioonireaktsioonides vabanevat energiat ei saa assimilatsiooniprotsessides otseselt kasutada. Esiteks muundatakse see energia ATP makroenergeetiliste sidemete energiaks ja alles seejärel kulutatakse orgaaniliste ühendite sünteesiks.
Kemolitoautotroofsed organismid:
Rauabakterid (Geobacter, Gallionella) oksüdeerivad kahevalentse raua raudrauaks.
Väävlibakterid (Desulfuromonas, Desulfobacter, Beggiatoa) oksüdeerivad vesiniksulfiidi molekulaarseks väävliks või väävelhappe sooladeks.
Nitrifitseerivad bakterid (Nitrobacteraceae, Nitrosomonas, Nitrosococcus) oksüdeerivad orgaanilise aine lagunemisel tekkinud ammoniaagi lämmastik- ja lämmastikhapeteks, mis koosmõjul mulla mineraalidega moodustavad nitriteid ja nitraate.
Tioonbakterid (Thiobacillus, Acidithiobacillus) on võimelised oksüdeerima tiosulfaate, sulfiteid, sulfiide ja molekulaarset väävlit väävelhappeks (sageli lahuse pH olulise langusega), oksüdatsiooniprotsess erineb väävlibakterite omast (eriti et tioonbakterid ei ladesta rakusisest väävlit). Mõned tioonsete bakterite esindajad on äärmuslikud atsidofiilid (võimelised ellu jääma ja paljunema, kui lahuse pH langeb 2-ni), taluvad suuri raskmetallide kontsentratsioone ning oksüdeerivad metallilist ja raudraudset rauda (Acidithiobacillus ferrooxidans) ning leostavad maakidest raskmetalle. .
Vesinikbakterid (Hydrogenophilus) on võimelised oksüdeerima molekulaarset vesinikku ja on mõõdukalt termofiilsed (kasvavad temperatuuril 50 °C)
Kemosünteetilised organismid (näiteks väävlibakterid) võivad elada ookeanides suurel sügavusel, kohtades, kus vesiniksulfiid väljub maakoore murdudest vette. Loomulikult ei suuda valguskvandid vette tungida umbes 3-4 kilomeetri sügavusele (sel sügavusel asub enamik ookeanilõhede tsoone). Seega on kemosünteetikumid ainsad organismid maa peal, mis ei sõltu päikesevalguse energiast.
Seevastu ammoniaak, mida kasutavad nitrifitseerivad bakterid, eraldub pinnasesse taimede või loomsete ainete mädanemisel. Sel juhul sõltub kemosünteetiliste ainete elutähtis aktiivsus kaudselt päikesevalgusest, kuna ammoniaak tekib päikeseenergiast saadavate orgaaniliste ühendite lagunemisel.
Kemosünteetiliste ainete roll kõigi elusolendite jaoks on väga suur, kuna need on asendamatuks lüliks kõige olulisemate elementide – väävli, lämmastiku, raua jne – loomulikus ringluses. Kemosünteetikumid on olulised ka selliste mürgiste ainete, nagu ammoniaak ja ammoniaak, looduslike tarbijatena. vesiniksulfiid. Suur tähtsus on nitrifitseerivatel bakteritel, kes rikastavad mulda nitritite ja nitraatidega – peamiselt nitraatide kujul omastavad taimed lämmastikku. Teatud kemosünteetilisi aineid (eelkõige väävlibaktereid) kasutatakse reovee puhastamiseks.
Kaasaegsete hinnangute kohaselt võib eelkõige merepõhja all asuva maa-aluse biosfääri biomass, mis sisaldab kemosünteetilisi anaeroobseid metaani oksüdeerivaid arhebaktereid, ületada ülejäänud biosfääri biomassi.
Meioos. Meioosi esimese ja teise jagunemise tunnused. Bioloogiline tähtsus. Erinevus meioosi ja mitoosi vahel
Arusaam, et sugurakud on haploidsed ja peavad seetõttu moodustuma spetsiaalse raku jagunemise mehhanismi abil, tuli vaatluste tulemusena, mis samuti viitasid peaaegu esimest korda, et kromosoomid sisaldavad geneetilist teavet. 1883. aastal avastati, et teatud tüüpi usside munaraku ja spermatosoidide tuumad sisaldavad ainult kahte kromosoomi, viljastatud munarakus aga juba neli. Kromosoomi teooria pärilikkus võib seega seletada pikaajalist paradoksi, et isa ja ema roll järglaste omaduste määramisel näib sageli olevat sama, hoolimata munaraku ja sperma suuruse tohutust erinevusest.
Selle avastuse teiseks oluliseks tagajärjeks oli see, et sugurakud peavad moodustuma eritüüpi tuumajagunemise tulemusena, mille käigus kogu kromosoomide komplekt jaguneb täpselt pooleks. Seda tüüpi jagunemist nimetatakse meioosiks (sõna Kreeka päritolu, mis tähendab "vähenemist". Teise rakkude jagunemise tüübi nimi - mitoos - pärineb kreeka sõnast, mis tähendab "niit", see nimevalik põhineb kromosoomide niidilaadsel välimusel nende kondenseerumisel tuuma jagunemise ajal - see protsess toimub nii mitoosi kui ka meioosi ajal; ) Kromosoomide käitumine meioosi ajal, kui nende arv väheneb, osutus arvatust keerukamaks. Seetõttu selgusid meiootilise jagunemise kõige olulisemad tunnused alles 30ndate alguseks tohutu hulga põhjalike uuringute tulemusena, mis ühendasid tsütoloogiat ja geneetikat.
Esimesel meiootilisel jagunemisel pärib iga tütarrakk kahest homoloogist kaks koopiat ja sisaldab seetõttu diploidset kogust DNA-d.
Haploidsete sugurakkude tuumade moodustumine toimub meioosi teise jagunemise tulemusena, mille käigus kromosoomid reastuvad uue spindli ekvaatorile ja ilma edasise DNA replikatsioonita eralduvad sõsarkromatiidid üksteisest, nagu tavalises mitoosis, moodustades haploidse DNA komplektiga rakud.
Seega koosneb meioos kahest raku jagunemisest, mis järgneb ühele kromosoomi dubleerimise faasile, nii et iga meioosi sisenev rakk annab neli haploidset rakku.
Mõnikord kulgeb meioosi protsess ebanormaalselt ja homoloogid ei saa üksteisest eralduda - seda nähtust nimetatakse kromosoomide mittedisjunktsiooniks. Mõned sel juhul moodustunud haploidsed rakud saavad ebapiisava arvu kromosoome, teised aga omandavad oma täiendavad koopiad. Sellistest sugurakkudest moodustuvad defektsed embrüod, millest enamik sureb.
Meioosi esimese jagunemise profaasis tekivad kromosoomide konjugatsiooni (sünapsi) ja eraldamise käigus neis keerulised morfoloogilised muutused. Vastavalt nendele muudatustele on profaas jagatud viieks järjestikuseks etapiks:
leptoteen;
sigoteen;
pahhüteen;
diploteen;
diakinees.
Kõige silmatorkavam nähtus on kromosoomide lähedane lähendamine sügoteenis, kui igas kahevalentsis õdekromatiidide paaride vahel hakkab moodustuma spetsiaalne struktuur, mida nimetatakse sünaptonemaalseks kompleksiks. Kromosoomide täieliku konjugeerumise hetke peetakse pahhüteeni alguseks, mis kestab tavaliselt mitu päeva pärast kromosoomide eraldumist, algab diploteeni staadium, mil chiasmata muutuvad esmakordselt nähtavaks.
Pärast pika I profaasi lõppu viivad kaks tuumajaotust ilma DNA sünteesi eraldusperioodita meioosi protsessi lõpuni. Need etapid ei võta tavaliselt rohkem kui 10% kogu meioosi jaoks vajalikust ajast ja neil on samad nimetused kui mitoosi vastavatel etappidel. Ülejäänud osa meioosi esimesest jagunemisest jaguneb metafaasiks I, anafaasiks I ja telofaasiks I. Esimese jagunemise lõpuks kromosoomikomplekt taandub, muutudes tetraploidsest diploidseks, nagu mitoosi puhul, ja moodustub kaks rakku. ühest rakust. Otsustav erinevus seisneb selles, et meioosi esimese jagunemise ajal saab iga rakk kaks tsentromeeriga ühendatud sõsarkromatiidi ja mitoosi käigus sisenevad kaks eraldatud kromatiidi.
Lisaks toimub pärast lühikest II interfaasi, milles kromosoomid ei kahekordistu, kiiresti teine jagunemine - II profaas, II anafaas ja II telofaas. Selle tulemusena moodustub igast meioosi sisenenud diploidsest rakust neli haploidset tuuma.
Meioos koosneb kahest järjestikusest rakujagunemisest, millest esimene kestab peaaegu sama kaua kui kogu meioos ja on palju keerulisem kui teine.
Pärast esimese meiootilise jagunemise lõppu moodustuvad kahes tütarrakus uuesti membraanid ja algab lühike interfaas. Sel ajal on kromosoomid mõnevõrra despiraliseeritud, kuid peagi need kondenseeruvad uuesti ja algab II profaas. Kuna sel perioodil DNA sünteesi ei toimu, näib, et mõnes organismis liiguvad kromosoomid otse ühest jagunemisest teise. Kõigi organismide II profaas on lühike: tuumaümbris hävib uue spindli moodustumisel, millele järgnevad kiires järjestuses metafaas II, anafaas II ja telofaas II. Nagu mitoosi puhul, moodustuvad kinetokoorfilamendid sõsarkromatiidides, mis ulatuvad tsentromeerist vastassuundadesse. Metafaasiplaadil hoitakse kahte õdekromatiidi koos kuni anafaasini, mil nad eralduvad kinetokooride äkilise eraldumise tõttu. Seega sarnaneb meioosi teine jagunemine tavalise mitoosiga, ainus oluline erinevus on see, et igast kromosoomist on üks koopia, mitte kaks, nagu mitoosi korral.
Meioos lõpeb tuumaümbriste moodustumisega II telofaasis moodustunud nelja haploidse tuuma ümber.
Üldiselt toodab meioosi ühest diploidsest rakust neli haploidset rakku. Gameotilise meioosi käigus moodustuvad tekkivatest haploidsetest rakkudest sugurakud. Seda tüüpi meioos on iseloomulik loomadele. Gameotiline meioos on tihedalt seotud gametogeneesi ja viljastumisega. Sügootilise ja eosmeioosi ajal tekitavad tekkivad haploidsed rakud eoseid või zoospoore. Seda tüüpi meioosid on iseloomulikud madalamatele eukarüootidele, seentele ja taimedele. Eoste meioos on tihedalt seotud sporogeneesiga. Seega on meioos sugulise ja mittesugulise (eoste) paljunemise tsütoloogiline alus.
Meioosi bioloogiline tähtsus on säilitada konstantne arv kromosoome seksuaalse protsessi juuresolekul. Lisaks toimub ületamise tulemusena rekombinatsioon - kromosoomides ilmnevad uued pärilike kalduvuste kombinatsioonid. Meioos annab ka kombinatiivse varieeruvuse – uute pärilike kalduvuste kombinatsioonide tekkimist edasise viljastamise käigus.
Meioosi kulgu juhib organismi genotüüp, suguhormoonide (loomadel), fütohormoonide (taimedel) ja paljude muude tegurite (näiteks temperatuur) kontrolli all.
Mõned organismid võivad teistele avaldada järgmist tüüpi mõjusid:
positiivne – üks organism saab kasu teise arvelt;
negatiivne - keha on kahjustatud millegi muu tõttu;
neutraalne - teine ei mõjuta keha kuidagi.
Seega on kahe organismi vaheliste suhete jaoks võimalikud järgmised võimalused sõltuvalt nende üksteisele avaldatavast mõjust:
Mutualism – looduslikes tingimustes ei saa populatsioonid üksteiseta eksisteerida (näide: seene ja vetikate sümbioos samblikus).
Protokoostöö - suhe on valikuline (näide: krabi ja anemooni suhe, anemone kaitseb krabi ja kasutab seda transpordivahendina).
Kommensalism – üks elanikkond saab suhtest kasu, teine aga ei saa kasu ega kahju.
Kooselu - üks organism kasutab teist (või oma kodu) elukohana, kahjustamata viimast.
Freeloading – üks organism toitub teise toidujääkidest.
Neutraalsus – mõlemad populatsioonid ei mõjuta teineteist kuidagi.
Amensalism, antibioos – üks populatsioon mõjutab negatiivselt teist, kuid ei koge ise negatiivset mõju.
Kisklus on nähtus, mille puhul üks organism toitub teise organismi organitest ja kudedest, ilma sümbiootilise suhteta.
Konkurents – mõlemad populatsioonid mõjutavad üksteist negatiivselt.
Loodus teab arvukalt näiteid sümbiootilistest suhetest, millest saavad kasu mõlemad partnerid. Näiteks liblikõieliste taimede ja mullabakterite Rhizobium sümbioos on looduses toimuva lämmastikuringe jaoks äärmiselt oluline. Need bakterid, mida nimetatakse ka lämmastikku siduvateks bakteriteks, settivad taimede juurtele ja neil on võime "kinnitada" lämmastikku, st lõhkuda tugevaid sidemeid atmosfääri vaba lämmastiku aatomite vahel, võimaldades lämmastikku siduda. taimele ligipääsetavad ühendid, näiteks ammoniaak. Sel juhul on mõlemapoolne kasu ilmne: juured on bakterite elupaigaks ja bakterid varustavad taime vajalike toitainetega.
Samuti on arvukalt näiteid sümbioosist, mis on kasulik ühele liigile ega too teisele liigile kasu ega kahju. Näiteks inimese soolestikus elab mitut tüüpi baktereid, mille esinemine on inimesele kahjutu. Samamoodi elavad bromeliaadideks kutsutud taimed (sealhulgas näiteks ananass) puuokstel, kuid saavad toitaineid õhust. Need taimed kasutavad puud toestamiseks ilma toitaineid ilma jätmata.
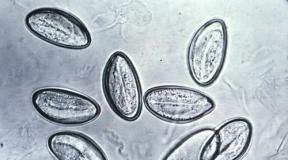
Lamedad ussid. Morfoloogia, süstemaatika, peamised esindajad. Arengutsüklid. Nakatumise teed. Ärahoidmine
Lamedad ussid on organismide rühm, mis enamikus kaasaegsetes klassifikatsioonides on varjupaiga auaste, mis ühendab suurt hulka primitiivseid ussilaadseid selgrootuid, kellel pole kehaõõnsust. Tänapäevasel kujul on rühm selgelt parafüleetiline, kuid praegune teadustöö ei võimalda välja töötada rahuldavat rangelt fülogeneetilist süsteemi ja seetõttu kasutavad zooloogid traditsiooniliselt seda nimetust.
Lameusside tuntumad esindajad on planaaria (Turbellaria: Tricladida), maksa- ja kassirest (trematood), veise-paeluss, sea-paeluss, laia paeluss, ehhinokokk (paeluss).
Praegu arutletakse niinimetatud soolestiku turbellarlaste (Acoela) süstemaatilise positsiooni üle, kuna 2003. aastal tehti ettepanek eristada nad iseseisvaks hõimkonnaks.
Keha on kahepoolselt sümmeetriline, selgelt piiritletud pea- ja sabaotstega, dorsoventraalses suunas mõnevõrra lapik, suurte esindajate korral tugevalt lame. Kehaõõnsus ei ole välja arenenud (v.a mõned paelusside ja liblikate elutsükli faasid). Gaaside vahetus toimub kogu keha pinnal; hingamiselundid ja veresooned puuduvad.
Keha väliskülg on kaetud ühekihilise epiteeliga. Ripsmelistel ussidel ehk turbellaritel koosneb epiteel ripsmeid kandvatest rakkudest. Lestadel, monogeenidel, tsestoodidel ja paelussidel puudub ripsepiteel suurema osa oma elust (kuigi ripsmelised rakud võivad esineda vastsete kujul); nende sisekest esindab nn tegument, mis mõnes rühmas kannab mikrovilli või kitiinkonkse. Lamedad ussid, millel on tegument, klassifitseeritakse neodermadeks.
Epiteeli all on lihasekott, mis koosneb mitmest lihasrakkude kihist, mis ei ole üksikuteks lihasteks diferentseerunud (teatavat diferentseerumist täheldatakse ainult neelu ja suguelundite piirkonnas). Välise lihaskihi rakud on orienteeritud risti, sisemise kihi rakud aga piki keha eesmist-tagamist telge. Välist kihti nimetatakse ringlihase kihiks ja sisemist kihti pikisuunaliseks lihaskihiks.
Kõigis rühmades, välja arvatud tsestoodid ja paelussid, on neelu, mis viib soolestikku või, nagu nn soolestiku turbellaritel, seede parenhüümi. Soolestik on pimesi suletud ja suhtleb keskkonnaga ainult suuava kaudu. On täheldatud, et mitmel suurel turbellarlasel on anaalpoorid (mõnikord mitu), kuid see on pigem erand kui reegel. Väikestes vormides on soolestik sirge, suurtel (planaria, flukes) võib see olla väga hargnenud. Neelu paikneb kõhupinnal, sageli keha keskosas või sellele lähemale, mõnes rühmas on see nihkunud ettepoole. Cestode-kujulistel ja paelussidel pole soolestikku.
Närvisüsteem nn ortogonaalne tüüp. Enamikul neist on kuus pikisuunalist tüve (kaks keha selja- ja ventraalsel küljel ning kaks külgedel), mis on ühendatud põikkommissuuridega. Koos ortogoniga on parenhüümi perifeersetes kihtides paiknev enam-vähem tihe närvipõimik. Mõnedel ripsmeliste usside kõige arhailisematel esindajatel on ainult närvipõimik.
Paljudel vormidel on välja arenenud lihtsad valgustundlikud ocellid, mis ei ole võimelised nägema objekti, samuti tasakaaluelundid (stagotsüstid), puutetundlikud rakud (sensilla) ja keemilised meeleelundid.
Osmoregulatsioon viiakse läbi protonefridia abil - hargnevad kanalid, mis ühenduvad üheks või kaheks erituskanaliks. Mürgiste ainevahetusproduktide vabanemine toimub kas protonefridia kaudu erituva vedelikuga või akumuleerudes spetsiaalsetesse parenhüümirakkudesse (atrotsüüdid), mis täidavad "hoiupungade" rolli.
Valdav enamus esindajatest on hermafrodiidid, välja arvatud verelibled (skistosoomid) - nad on kahekojalised. Fluke munad on helekollase kuni tumepruuni värvusega ja nende ühel poolustel on kork. Uurimise käigus leitakse mune kaksteistsõrmiksoole sisust, väljaheitest, uriinist ja rögast.
Lestlaste esimeseks vaheperemeheks on erinevad molluskid, teiseks peremeheks kalad ja kahepaiksed. Lõplikud peremehed on erinevad selgroogsed.
Elutsükkel (polüsuu näitel) on ülimalt lihtne: munast väljub kalast lahkuv vastne, kes mõne aja pärast jälle kala külge kinnitub ja muutub täiskasvanud ussiks. Flukes on keerulisem arendustsükkel, vahetades 2-3 peremeest.
Genotüüp. Genoom. Fenotüüp. Fenotüübi arengut määravad tegurid. Domineerimine ja retsessiivsus. Geenide koostoime tunnuste määramisel: domineerimine, vahepealne manifestatsioon, kadominantsus
Genotüüp on antud organismi geenide kogum, mis erinevalt genoomi ja genofondi kontseptsioonidest iseloomustab indiviidi, mitte liiki (teine erinevus genotüübi ja genoomi vahel on see, et mõistesse "genoom" lisatakse ka mitte liike. -kodeerivad järjestused, mis ei kuulu „genotüübi“ mõiste alla). Koos keskkonnateguritega määrab see organismi fenotüübi.
Tavaliselt räägitakse genotüübist konkreetse geeni kontekstis polüploidsetel indiviididel, see tähistab antud geeni alleelide kombinatsiooni. Enamik geene esineb organismi fenotüübis, kuid fenotüüp ja genotüüp erinevad järgmistes aspektides:
1. Vastavalt teabeallikale (genotüüp määratakse isendi DNA-d uurides, fenotüüp registreeritakse organismi välimust jälgides).
2. Genotüüp ei vasta alati samale fenotüübile. Mõned geenid ilmuvad fenotüübis ainult teatud tingimustel. Teisest küljest on mõned fenotüübid, näiteks loomade karvavärv, mitme geeni koostoime tulemus.
Genoom – organismi kõigi geenide kogum; selle täielik kromosoomikomplekt.
On teada, et DNA, mis on enamikus organismides geneetilise teabe kandja ja seega genoomi aluseks, ei hõlma ainult geene selle sõna tänapäevases tähenduses. Enamikku eukarüootsete rakkude DNA-st esindavad mittekodeerivad ("liigsed") nukleotiidjärjestused, mis ei sisalda teavet valkude ja RNA kohta.
Järelikult mõistetakse organismi genoomi all haploidse kromosoomikomplekti ja iga kromosoomivälise geneetilise elemendi kogu DNA-d, mis sisalduvad mitmerakulise organismi iduliini üksikus rakus. Erinevate liikide organismide genoomide suurused erinevad üksteisest oluliselt ning sageli puudub seos bioloogilise liigi evolutsioonilise keerukuse taseme ja selle genoomi suuruse vahel.
Fenotüüp on teatud arenguetapis indiviidile omaste omaduste kogum. Fenotüüp kujuneb genotüübi alusel, mida vahendavad mitmed keskkonnategurid. Diploidsetes organismides esinevad fenotüübis domineerivad geenid.
Fenotüüp on organismi väliste ja sisemiste omaduste kogum, mis on omandatud ontogeneesi (individuaalse arengu) tulemusena.
Vaatamata näiliselt rangele määratlusele on fenotüübi mõistel teatud ebakindlus. Esiteks ei ole enamik geneetilise materjali poolt kodeeritud molekule ja struktuure organismi välisilmes nähtavad, kuigi need on osa fenotüübist. Näiteks inimese veregrupid. Seetõttu peaks fenotüübi laiendatud määratlus hõlmama omadusi, mida saab tuvastada tehniliste, meditsiiniliste või diagnostiliste protseduuridega. Veel üks radikaalsem laiendus võib hõlmata õpitud käitumist või isegi organismi mõju keskkonnale ja teistele organismidele.
Fenotüüpi võib defineerida kui geneetilise informatsiooni "viimist" keskkonnategurite suunas. Esimeseks lähenduseks saame rääkida kahest fenotüübi tunnusest: a) eemaldamise suundade arv iseloomustab nende keskkonnategurite hulka, mille suhtes fenotüüp on tundlik – fenotüübi dimensiooni; b) eemaldamise "kaugus" iseloomustab fenotüübi tundlikkuse astet antud keskkonnateguri suhtes. Need omadused koos määravad fenotüübi rikkuse ja arengu. Mida mitmemõõtmelisem ja tundlikum on fenotüüp, mida kaugemal on fenotüüp genotüübist, seda rikkam see on. Kui võrrelda viirust, bakterit, ascarist, konna ja inimest, siis fenotüübi rikkus selles seerias suureneb.
Mõned fenotüübi omadused on otseselt määratud genotüübiga, näiteks silmade värv. Teised sõltuvad suuresti organismi vastasmõjust keskkonnaga – näiteks võivad identsed kaksikud erineda pikkuse, kaalu ja muude põhiliste füüsiliste omaduste poolest, hoolimata sellest, et nad kannavad samu geene.
Fenotüübiline dispersioon (määratud genotüübi dispersiooniga) on loodusliku valiku ja evolutsiooni põhieeldus. Organism tervikuna jätab (või ei jäta) järglasi, seega looduslik valik mõjutab populatsiooni geneetilist struktuuri kaudselt fenotüüpide panuse kaudu. Ilma erinevate fenotüüpideta pole evolutsiooni. Samal ajal ei kajastu retsessiivsed alleelid alati fenotüübi omadustes, vaid need säilivad ja võivad järglastele edasi kanduda.
Faktorid, millest sõltuvad fenotüübiline mitmekesisus, geneetiline programm (genotüüp), keskkonnatingimused ja juhuslike muutuste (mutatsioonide) sagedus, on kokku võetud järgmises seoses:
genotüüp + väliskeskkond + juhuslikud muutused → fenotüüp.
Genotüübi võimet moodustada ontogeneesis erinevaid fenotüüpe, olenevalt keskkonnatingimustest, nimetatakse reaktsiooninormiks. See iseloomustab keskkonna osalust tunnuse rakendamisel. Mida laiem on reaktsiooninorm, seda suurem on keskkonna mõju ja seda väiksem on genotüübi mõju ontogeneesis. Tavaliselt, mida mitmekesisemad on liigi elupaigatingimused, seda laiem on tema reaktsiooninorm.
Jätkamine
--PAGE_BREAK--
Dominantsus (dominantsus) on ühe geeni alleelide vahelise suhte vorm, milles üks neist (dominantne) surub (maskib) teise (retsessiivne) avaldumist ja määrab seega tunnuse avaldumise nii domineerivates homosügootides kui ka heterosügootides. .
Täieliku domineerimise korral ei erine heterosügoodi fenotüüp domineeriva homosügoodi fenotüübist. Ilmselt sisse puhtal kujul täielik domineerimine on äärmiselt haruldane või seda ei esine üldse.
Mittetäieliku domineerimise korral on heterosügootidel fenotüüp, mis jääb domineeriva ja retsessiivse homosügoodi fenotüüpide vahele. Näiteks kui ristuda puhtad snapdraakoni ja paljude teiste lillade ja valgete õitega lilletaimede liigid, on esimese põlvkonna isenditel roosad õied. Molekulaarsel tasandil võib mittetäieliku domineerimise lihtsaim seletus olla vaid ensüümi või muu valgu aktiivsuse kahekordne vähenemine (kui domineeriv alleel toodab funktsionaalset valku ja retsessiivne alleel defektset). Võib esineda ka teisi mittetäieliku domineerimise mehhanisme.
Mittetäieliku domineerimise korral on sama jagunemine genotüübi ja fenotüübi järgi vahekorras 1: 2: 1.
Koosdominantsiga, erinevalt mittetäielikust domineerimisest, ilmnevad heterosügootides tunnused, mille eest iga alleel vastutab, samaaegselt (segatuna). Tüüpiline näide kodominantsus – ABO veregruppide pärand inimestel. Kõigil AA (teine rühm) ja BB (kolmas rühm) genotüübiga inimeste järglastel on AB genotüüp (neljas rühm). Nende fenotüüp ei ole nende vanemate fenotüüpide vahepealne, kuna erütrotsüütide pinnal on mõlemad aglutinogeenid (A ja B). Kodominantsi ilmnemisel on võimatu nimetada ühte alleeli domineerivaks ja teist retsessiivseks, need mõisted kaotavad oma tähenduse: mõlemad alleelid mõjutavad fenotüüpi võrdselt. RNA ja valgu geeniproduktide tasandil ilmneb, et valdav enamus alleelsete geenide interaktsiooni juhtudest on kodominantsus, kuna heterosügootides kodeerib kumbki alleel tavaliselt RNA-d ja/või valguprodukti ning mõlemad valgud või RNA on kehas olemas.
Keskkonnategurid, nende koosmõju
Keskkonnategur on keskkonnaseisund, mis mõjutab organismi. Keskkonda kuuluvad kõik kehad ja nähtused, millega organism on otseses või kaudses suhtes.
Samal keskkonnateguril on kooselusorganismide elus erinev tähendus. Näiteks mulla soolarežiim mängib taimede mineraalse toitumise juures esmast rolli, kuid on ükskõikne enamiku maismaaloomade suhtes. Valgustuse intensiivsus ja valguse spektraalne koostis on fototroofsete taimede elus äärmiselt olulised ning heterotroofsete organismide (seente ja veeloomade) elus ei avalda valgus nende elutegevusele märgatavat mõju.
Keskkonnategurid mõjutavad organisme erineval viisil. Need võivad toimida ärritajatena, mis põhjustavad adaptiivseid muutusi füsioloogilistes funktsioonides; piirajatena, mis muudavad teatud organismide eksisteerimise antud tingimustes võimatuks; kui modifikaatorid, mis määravad organismide morfoloogilisi ja anatoomilisi muutusi.
Tavapärane on eristada biootilisi, antropogeenseid ja abiootilisi keskkonnategureid.
Biootilised tegurid on elusorganismide tegevusega seotud keskkonnategurite kogum. Nende hulka kuuluvad fütogeensed (taimed), zoogeensed (loomad), mikrobiogeensed (mikroorganismid) tegurid.
Antropogeensed tegurid on kõik paljud inimtegevusega seotud tegurid. Nende hulka kuuluvad füüsikalised (tuumaenergia kasutamine, reisimine rongides ja lennukites, müra ja vibratsiooni mõju jne), keemiline (mineraalväetiste ja pestitsiidide kasutamine, maakera reostus tööstus- ja transpordijäätmetega, suitsetamine, alkoholi ja narkootikumide tarbimine, ravimite liigtarbimine), bioloogilised (toit; organismid, millele inimene võib olla elupaigaks või toitumisallikaks), sotsiaalsed (seotud inimeste ja eluga ühiskonnas).
Abiootilised tegurid on kõik paljud tegurid, mis on seotud elutu looduse protsessidega. Nende hulka kuuluvad klimaatilised (temperatuur, niiskus, rõhk), edafogeensed (mehaaniline koostis, õhu läbilaskvus, pinnase tihedus), orograafilised (reljeef, kõrgus merepinnast), keemilised (õhu gaasiline koostis, vee soolane koostis, kontsentratsioon, happesus), füüsikalised (müra, magnetväljad, soojusjuhtivus, radioaktiivsus, kosmiline kiirgus).
Kui keskkonnategurid toimivad iseseisvalt, piisab keskkonnategurite kompleksi koosmõju kindlaksmääramiseks antud organismile "piirava teguri" mõistest. Reaalsetes tingimustes võivad keskkonnategurid aga üksteise mõju tugevdada või nõrgendada.
Keskkonnategurite koosmõju arvestamine on oluline teaduslik probleem. Eristada saab kolme peamist tegurite koostoime tüüpi:
aditiivne - tegurite koostoime on iga teguri mõjude lihtne algebraline summa iseseisvalt toimides;
sünergiline - tegurite ühine toime suurendab efekti (see tähendab, et nende koos toimimise mõju on suurem kui iga teguri mõjude lihtne summa, kui nad tegutsevad iseseisvalt);
antagonistlik - tegurite ühine toime nõrgendab mõju (st nende ühistegevuse mõju on väiksem kui iga teguri mõjude lihtsumma).
Kasutatud kirjanduse loetelu
Gilbert S. Arengubioloogia. - M., 1993.
Green N., Stout W., Taylor D. Bioloogia. - M., 1993.
Nebel B. Keskkonnateadus. - M., 1993.
Carroll R. Selgroogsete paleontoloogia ja evolutsioon. - M., 1993.
Leninger A. Biokeemia. - M., 1974.
Slyusarev A.A. Bioloogia üldgeneetikaga. - M., 1979.
Watson D. Geeni molekulaarbioloogia. - M., 1978.
Chebyshev N.V., Supryaga A.M. Algloomad. - M., 1992.
Tšebõšev N.V., Kuznetsov S.V. Rakubioloogia. - M., 1992.
Yarygin V.N. Bioloogia. - M., 1997.
Sisu Mikroskoop Nimed, mis mängisid rolli rakkude uurimisel Rakuteooria põhiprintsiibid Rakustruktuurid: Rakuorganellid: Rakumembraan Tsütoplasma Tuum Ribosoomid Golgi kompleks ER Lüsosoomid MitokondridMitokondrid Plastiidid Rakukeskus Liikumisorganellid

Mikroskoop Anton Van Leeuwenhoek Anton Van Leeuwenhoek lõi maailma esimese mikroskoobi, mis võimaldas uurida raku mikrostruktuuri. Mikroskoobi täiustamisega avastasid teadlased rakust üha rohkem tundmatuid osi, elutähtsaid protsesse, mida oli võimalik jälgida valgusmikroskoobis. Riis. 1: Leeuwenhoeki mikroskoop Kahekümnendal sajandil leiutatud elektrimikroskoop ja selle täiustatud mudel võimaldavad näha rakustruktuuride mikroskoopilist struktuuri. Mahulise skaneerimisega saate näha raku ja selle organellide struktuuri sellisena, nagu need on oma loomulikus keskkonnas, elusorganismis. Riis. 2: elektrimikroskoop

Nimed, mis mängisid rolli rakkude uurimisel Anton van Leeuwenhoek Anton van Leeuwenhoek oli esimene, kes uuris üherakulisi organisme mikroskoobi all. Robert Hooke Robert Hooke pakkus välja termini "Cell". T. Schwann T. Schwann ja M. Schleiden – sõnastasid rakuteooria 19. sajandi keskel.M. Schleideni rakuteooria R. Brown R. Brown – sisse XIX algus sajandil nägin leherakkude sees tihedat moodustist, mida nimetasin tuumaks. R. Virchow R. Virchow – tõestas, et rakud on võimelised jagunema ja pakkus välja lisandi rakuteooriale.

Rakuteooria põhisätted 1. Kõik elusolendid, ainuraksete kuni suurte taime- ja loomaorganismideni, koosnevad rakkudest. 2. Kõik rakud on ehituselt, keemiliselt koostiselt ja elutähtsatelt funktsioonidelt sarnased. 3. Rakud on spetsialiseerunud ja mitmerakuliste organismide koostise ja funktsioonide poolest ning on võimelised iseseisvaks eluks. 4. Rakud on valmistatud rakkudest. Rakk on aluseks emaraku lagunemisele kaheks tütarrakuks.

Rakustruktuurid Rakumembraan Enamiku organellide seinad moodustab rakumembraan. Rakumembraani struktuur: see on kolmekihiline. Paksus - 8 nanomeetrit. 2 kihti moodustavad lipiide, mis sisaldavad valke. Membraanvalgud moodustavad sageli membraanikanaleid, mille kaudu transporditakse kaaliumi, kaltsiumi ja naatriumi ioone. Suured valkude, rasvade ja süsivesikute molekulid sisenevad rakku fagotsütoosi ja pinotsütoosi abil. Fagotsütoos on rakumembraaniga ümbritsetud tahkete osakeste sisenemine raku tsütoplasmasse. Pinotsütoos on rakumembraaniga ümbritsetud vedelate tilkade sisenemine raku tsütoplasmasse. Ainete vool läbi membraani toimub valikuliselt, lisaks piirab see rakku, eraldab selle teistest, keskkonnast, annab kuju ja kaitseb kahjustuste eest. Riis. 4: A – fagotsütoosi protsess; B – pinotsütoosi protsess Joon. 3: rakumembraani struktuur

Rakkude struktuurid Tsütoplasma. Tuum. Tsütoplasma on raku poolvedel sisaldus, mis sisaldab kõiki raku organelle. Kompositsioon sisaldab erinevaid orgaanilisi ja anorgaanilisi aineid, vett ja sooli. Tuum: Ümmargune, tihe, tume keha taimede, seente ja loomade rakkudes. Ümbritsetud tuumamembraaniga. Membraani välimine kiht on kare, sisemine kiht sile. Paksus - 30 nanomeetrit. Omab poore. Tuuma sees on tuumamahl. Sisaldab kromatiini niite. Kromatiin - DNA + VALK. Jagamise ajal keritakse DNA valgule, nagu pool. Nii tekivad kromosoomid. Inimestel on keha somaatilistel rakkudel 46 kromosoomi. See on diploidne (täielik, topelt) kromosoomide komplekt. Sugurakkudel on 23 kromosoomi (haploidne, pool). Liigispetsiifilist kromosoomide komplekti rakus nimetatakse karüotüübiks. Organismid, mille rakkudel puudub tuum, nimetatakse prokarüootideks. Eukarüootid on organismid, mille rakud sisaldavad tuuma. Riis. 6: Meeste kromosoomikomplekt Joonis fig. 5: Põhistruktuur

Rakuorganellid Ribosoomid Organellid on sfäärilise kujuga, läbimõõduga nanomeetrid. Need koosnevad DNA-st ja valgust. Ribosoomid moodustuvad tuuma tuumades ja sisenevad seejärel tsütoplasmasse, kus nad hakkavad täitma oma funktsiooni - valgusünteesi. Tsütoplasmas paiknevad ribosoomid kõige sagedamini krobelisel endoplasmaatilisel retikulumil. Harvemini hõljuvad nad vabalt raku tsütoplasmas. Riis. 7: Eukarüootse raku ribosoomi struktuur

Rakuorganellid Golgi kompleks Need on õõnsused, mille seinad on moodustatud ühest membraanikihist, mis paiknevad virnadena tuuma lähedal. Sees on sünteesitud ained, mis kogunevad rakus. Vesiikulid vabanevad Golgi kompleksist ja moodustuvad lüsosoomideks. Riis. 8: Golgi aparaadi ehitusskeem ja mikrograaf

ER-raku organellid EPS on endoplasmaatiline retikulum. See on tuubulite võrgustik, mille seinad moodustab rakumembraan. Torude paksus on 50 nanomeetrit. EPS on kahte tüüpi: sile ja granuleeritud (kare). Sile täidab transpordifunktsiooni, kare aga (selle pinnal ribosoomid) sünteesib valke. Riis. 9: Granuleeritud EPS-i lõigu elektronmikroskoopia

Rakuorganellid Lüsosoomid Lüsosoom on väike vesiikul, mille läbimõõt on vaid 0,5–1,0 mikronit ja mis sisaldab suurt hulka ensüüme, mis võivad hävitada toiduaineid. Üks lüsosoom võib sisaldada 30–50 erinevat ensüümi. Lüsosoomid on ümbritsetud membraaniga, mis talub nende ensüümide toimet. Lüsosoomid moodustuvad Golgi kompleksis. Riis. 10: diagramm toiduosakeste seedimisest rakus lüsosoomi abil

Rakuorganellid Mitokondrid Mitokondrite ehitus: Ümarad, ovaalsed, vardakujulised kehad. Pikkus -10 mikromeetrit, läbimõõt -1 mikromeeter. Seinad on moodustatud kahest membraanist. Välimine on sile, sisemine eenditega - cristae. Sisemine osa on täidetud ainega, mis sisaldab suurt hulka ensüüme, DNA-d, RNA-d. Seda ainet nimetatakse maatriksiks. Funktsioonid: Mitokondrid toodavad ATP molekule. Nende süntees toimub kristallidel. Enamik mitokondreid leidub lihasrakkudes. Riis. 11: Mitokondrite struktuur

Rakuorganellid Plastiidid Plastiide on kolme tüüpi: leukoplastid – värvitud, kloroplastid – rohelised (klorofüll), kromoplastid – punased, kollased, oranžid. Plastiide leidub ainult taimerakkudes. Kloroplastid on sojaoa tera kujuga. Seinad on moodustatud kahest membraanist. Väliskiht on sile, sisemisel kihil on väljaulatuvad osad ja voltid, mis moodustavad mullide virnad, mida nimetatakse granaks. Grana sisaldab klorofülli, sest kloroplastide põhiülesanne on fotosüntees, mille tulemusena tekivad süsihappegaasist ja veest süsivesikud ja ATP. Kloroplastide sees on DNA, RNA, ribosoomide ja ensüümide molekulid. Nad võivad ka jagada (korrutada). Riis. 12: Kloroplasti struktuur

Rakuorganellid Rakukeskus Madalamatel taimedel ja loomadel on tuuma lähedal kaks sentiooli, see on rakukeskus. Need on kaks silindrilist keha, mis asuvad üksteisega risti. Nende seinad on moodustatud 9 mikrotuubulite kolmikust. Mikrotuubulid moodustavad raku tsütoskeleti, mida mööda liiguvad organellid. Jagunemise ajal moodustab rakukeskus spindli filamente, samal ajal kui see kahekordistub, ühele poolusele läheb 2 tsentriooli ja 2 teisele. Riis. 13: A – struktuurskeem ja B – tsentriooli elektronmikrofoto

Rakuorganellid Liikumisorganellid Liikumisorganellid on ripsmed ja lipukesed. Ripsmed on lühemad - neid on rohkem ja lipukesed on pikemad - neid on vähem. Need on moodustatud membraanist ja sisaldavad nende sees mikrotuubuleid. Mõnel liikumisorganellil on basaalkehad, mis kinnitavad need tsütoplasmasse. Liikumine toimub torude üksteise kohal libisemise tõttu. Inimese hingamisteedes on ripsepiteelil ripsmed, mis väljutavad tolmu, mikroorganisme ja lima. Algloomadel on lipud ja ripsmed. Riis. 14: Liikumisvõimelised üherakulised organismid


Anton van Leeuwenhoek Ta sündis 24. oktoobril 1632 Hollandis Delfti linnas. Tema sugulased olid lugupeetud linnakodanikud ning tegelesid korvipunumise ja õlle valmistamisega. Leeuwenhoeki isa suri varakult ja ema saatis poisi kooli, unistades temast ametnikuks saada. Kuid 15-aastaselt jättis Anthony kooli pooleli ja läks Amsterdami, kus hakkas õppima riidepoes kaubandust, töötades seal raamatupidaja ja kassapidajana. 21-aastaselt naasis Leeuwenhoek Delfti, abiellus ja avas oma tekstiiliäri. Tema elust on järgmise 20 aasta jooksul teada väga vähe, välja arvatud see, et tal oli mitu last, kellest enamik suri, ja et ta abiellus teist korda. Samuti on teada, et ta sai valvuri ametikoha kohalikus raekojas asuv kohtukoda, mis tänapäevaste ideede järgi vastab kojamehe, koristaja ja kergitaja kombinatsioonile ühes isikus. Leeuwenhoekil oli oma hobi. Töölt tulles lukustas ta end oma kabinetti, kuhu tollal isegi abikaasat ei lastud, ja uuris entusiastlikult suurendusklaaside all mitmesuguseid esemeid. Kahjuks neid prille liiga palju ei suurendatud. Seejärel proovis Leeuwenhoek teha oma mikroskoobi lihvklaasist, mis tal ka õnnestus.

Robert Hooke (ing. Robert Hooke; Robert Hook, 18. juuli 1635, Isle of Wight, 3. märts 1703 London) inglise loodusteadlane, entsüklopedist. Hooke’i pastorist isa valmistas ta algul ette vaimulikuks tegevuseks, kuid poisi kehva tervise ja ülesnäidatud mehaanikaga tegelemise võime tõttu määras ta kellassepa erialale. Hiljem aga hakkas noor Hooke huvi tundma teaduslike õpingute vastu ja selle tulemusena suunati ta Westminsteri kooli, kus ta õppis edukalt ladina, vanakreeka ja heebrea keelt, kuid tundis erilist huvi matemaatika vastu ning näitas üles suurepärast võimekust füüsikas ja leiutistes. mehaanika. Tema võimet õppida füüsikat ja keemiat tunnustasid ja hindasid teadlased Oxfordi ülikoolis, kus ta asus õppima 1653. aastal; Esmalt sai temast keemik Willise ja seejärel kuulsa Boyle'i assistent. Oma 68-aastase elu jooksul oli Robert Hooke vaatamata kehvale tervisele õpingutes väsimatu ning tegi palju teaduslikke avastusi, leiutisi ja täiustusi. 1663. aastal võttis Londoni Kuninglik Selts tema avastuste kasulikkust ja tähtsust tunnistades ta liikmeks; seejärel määrati ta Greshami kolledži geomeetriaprofessoriks.

Robert Hooke'i avastused Hooke'i avastused hõlmavad järgmist: proportsionaalsuse avastamine elastse venitamise, kokkusurumise ja painutamise ning neid tekitavate pingete vahel, universaalse gravitatsiooniseaduse mõningane esialgne sõnastus (Newton vaidlustas Hooke'i prioriteedi, kuid ilmselt mitte esialgne koostis), õhukeste plaatide värvide avastamine, jää sulamistemperatuuri ja vee keemise püsivus, valguse lainelise leviku idee ja gravitatsiooni idee, a. elusrakk (kasutades mikroskoopi, mida ta täiustas; Hooke ise omab mõistet "rakk" - inglise rakk) ja palju muud. Esiteks tuleks öelda kella liikumise reguleerimiseks mõeldud spiraalvedru kohta; selle leiutise tegi ta aastatel 1656–1666 leiutas ta vesiloodi, 1665. aastal esitas ta Kuninglikule Seltsile väikese kvadrandi, milles alidaadi liigutati mikromeetri kruvi abil, nii et oli võimalik lugeda minuteid. ja sekundid; edasi, kui leiti, et on mugav asendada astronoomiliste instrumentide dioptrid torudega, tegi ta ettepaneku asetada okulaari niitvõrk. Lisaks leiutas ta optilise telegraafi, miinimumtermomeetri ja salvestava vihmamõõturi; tegi vaatlusi, et teha kindlaks maakera pöörlemise mõju kehade kukkumisele ja uuris paljusid Joon. 3: Hooke'i mikroskoop füüsikaliste küsimustega, näiteks karvasuse, agregatsiooni, õhu kaalumise, jää erikaalu mõjude kohta, leiutas spetsiaalse hüdromeetri jõevee värskusastme (water-poise) määramiseks. Aastal 1666 esitles Hooke Royal Societyle enda leiutatud spiraalsete hammasrataste mudelit, mida ta kirjeldas hiljem raamatus Lectiones Cutlerianae (1674).

T. Schwann Theodor Schwann () sündis 7. detsembril 1810. aastal Düsseldorfi lähedal Reini-äärses Neussis, õppis Kölnis jesuiitide gümnaasiumis, õppis 1829. aastast arstiteadust Bonnis, Warzburgis ja Berliinis. Ta sai doktorikraadi 1834. aastal ja avastas pepsiini 1836. aastal. Schwanni monograafia “Microscopic Studies on the Sarnasus loomade ja taimede struktuuris ja kasvus” (1839) tõi talle ülemaailmse kuulsuse. Aastast 1839 oli ta anatoomiaprofessor Belgias Leuvenis ja 1848. aastast Lüttichis. Schwann oli vallaline ja oli usklik katoliiklane. Ta suri Kölnis 11. jaanuaril 1882. aastal. Tema väitekiri vajadusest atmosfääriõhk kana arenguks (1834) tutvustas õhu rolli organismide arenguprotsessides. Hapniku vajadust fermentatsiooniks ja mädanemiseks demonstreerisid ka Gay-Lussaci katsed. Schwanni tähelepanekud taaselustasid huvi spontaanse genereerimise teooria vastu ja taaselustasid idee, et kuumutamise tõttu kaotab õhk elujõu, mis on vajalik elusolendite tekkeks. Schwann püüdis tõestada, et kuumutatud õhk ei sega eluprotsessi. Ta näitas, et konn hingab soojas õhus normaalselt. Kui aga kuumutatud õhk lastakse läbi pärmisuspensiooni, millele on lisatud suhkrut, siis käärimist ei toimu, kusjuures kuumutamata pärm areneb kiiresti. Schwann jõudis oma kuulsate veini kääritamise katseteni teoreetiliste ja filosoofiliste kaalutluste põhjal. Ta kinnitas mõtet, et veini käärimist põhjustavad elusorganismid – pärm. Schwanni kuulsaimad tööd on histoloogia vallas, aga ka rakuteooriale pühendatud teosed. Olles tutvunud M. Schleideni töödega, vaatas Schwann läbi kogu tol ajal kättesaadava histoloogilise materjali ja leidis põhimõtte, kuidas võrrelda taimerakke ja loomade elementaarseid mikroskoopilisi struktuure. Võttes tuuma kui rakustruktuuri iseloomulikku elementi, suutis Schwann tõestada taime- ja loomarakkude ühist struktuuri. 1839. aastal ilmus Schwanni klassikaline teos "Microscopic Studies on the Correspondence in the Structure and Growth of Animals and Plants".

M. Schleiden Schleiden Matthias Jacob (, Hamburg - , Frankfurt Maini ääres), saksa botaanik. Ta õppis õigusteadust Heidelbergis, botaanikat ja meditsiini Göttingeni, Berliini ja Jena ülikoolides. Jena ülikooli botaanikaprofessor (1839–62), aastast 1863 - antropoloogiaprofessor Dorpati ülikoolis (Tartu). Teadusliku uurimistöö põhisuunaks on tsütoloogia ja taimefüsioloogia. 1837. aastal pakkus Schleiden välja uue taimerakkude moodustumise teooria, mis põhines ideel raku tuuma otsustavast rollist selles protsessis. Teadlane uskus, et uus rakk puhuti justkui tuumast välja ja seejärel kaeti rakuseinaga. Schleideni uurimistöö aitas kaasa T. Schwanni rakuteooria loomisele. Tuntud on Schleideni töid kõrgemate taimede rakustruktuuride arengu ja diferentseerumise kohta. 1842. aastal avastas ta esimest korda tuumast nukleoolid. Teadlase kuulsaimate tööde hulgas on "Botaanika alused" (Grundz ge der Botanik, 1842–1843)

R. Brown Robert Brown (sündinud 21. detsember 1773, Montrose – 10. juuni 1856) oli silmapaistev inglise botaanik. Sündis 21. detsembril Šotimaal Montorosas, õppis Aberdeenis ja Edinburghis ning 1795. a. astus lipnikuna ja abikirurgina Šoti miilitsa rügementi, millega ta viibis Iirimaal. Tema innukas loodusteaduste õppimine tõi talle sõpruse Sir Joseph Bankiga, kelle soovitusel määrati ta botaanikuks 1801. aastal kapten Flinderi juhtimisel Austraalia rannikut uurima. Koos kunstnik Ferdinand Baueriga külastas ta mõnda Austraaliat, seejärel Tasmaaniat ja Bassi väina saari. 1805. aastal naasis Brown Inglismaale, tuues endaga kaasa umbes 4000 Austraalia taimeliiki; ta töötas mitu aastat selle rikkaliku materjali väljatöötamisega, mida keegi polnud kunagi kaugetest riikidest toonud. Oma kalli looduslooliste tööde kogu raamatukoguhoidja Sir Banki tehtud Brown avaldas: "Prodromus florae Novae Hollandiae" (London, 1810), mille Oken trükkis "Isises" ja Nees von Esenbeck (Nürnberg, 1827) koos lisadega. . See eeskujulik töö andis taimegeograafiale (fütogeograafiale) uue suuna. Samuti koostas ta polaarmaadele reisijate Rossi, Parry ja Clappertoni aruannetes botaanika lõike, aitas kirurg Richardsoni, kes kogus Frankliniga reisil palju huvitavat; järk-järgult kirjeldas herbaariume, mida kogus: Gorsfield Javas aastatel. Oudney ja Clapperton Kesk-Aafrikas, Christian Smith, Tuquay kaaslane ekspeditsioonil mööda Kongot. Looduslik süsteem võlgneb talle palju: ta püüdles võimalikult suure lihtsuse poole nii liigituses kui ka terminoloogias, vältis igasuguseid tarbetuid uuendusi; tegi palju vanade määratluste parandamiseks ja uute perekondade loomiseks. Ta töötas ka taimefüsioloogia alal: uuris tolmuka arengut ja plasmakehade liikumist selles.

R. Virchow () (saksa: Rudolf Ludwig Karl Virchow) 19. sajandi teise poole saksa teadlane ja poliitik, rakuteooria rajaja bioloogias ja meditsiinis; oli tuntud ka arheoloogina. Ta sündis 13. oktoobril 1821 Schiefelbeini linnas Preisimaa Pommeri provintsis. Pärast kursuse lõpetamist Berliini Friedrich-Wilhelmi Meditsiiniinstituudis 1843. aastal sai V.-st esmalt assistent ja seejärel dissektor Berliini Charité haiglas. 1847. aastal sai ta õpetamisõiguse ja asutas koos Benno Reinhardiga (1852) ajakirja “Archiv für pathol. Anatoomia u. Füsioloogia u. kliinikusse. Medicine”, mis on nüüdseks tuntud kogu maailmas Virchow arhiivi nime all. 1848. aasta alguses saadeti Virchow Ülem-Sileesiasse uurima seal valitsenud näljatüüfuse epideemiat. Tema Arhiivis avaldatud ja suurt teaduslikku huvi pakkuv aruanne selle reisi kohta on samal ajal värvitud poliitilistest ideedest 1848. aasta vaimus. See asjaolu, nagu ka tema üldine osalemine tolleaegsetes reformiliikumistes, tekitas Preisi valitsuses tema vastumeelsuse ja ajendas teda vastu võtma Würzburgi ülikoolis talle pakutud tavalist patoloogilise anatoomia õppetooli, mis kiiresti tema nime ülistas. Aastal 1856 naasis ta Berliini patoloogilise anatoomia, üldpatoloogia ja teraapia professorina ning vastloodud patoloogilise instituudi direktorina, kuhu jäi kuni oma elu lõpuni. Eriti palju võlgnevad Venemaa arstiteadlased Virchowile ja tema instituudile.

MBOU Jasnogorski keskkool
Bioloogia
10 A klass
Õpik
Teema:
Sihtmärk:
Ülesanded:
Varustus:
Tundide ajal:
Slaid 1
1.
Vestlus probleemide üle (slaid nr 2)
1. Mis on noosfäär?
2. Uue materjali õppimine
Tunniplaan:
3. Struktuurielemendid.
4.Põhiprotsessid.
5. Organisatsiooni tunnused.
3. Konsolideerimine
Õpetaja teeb kokkuvõtte:
Küsimused
D/z. par.13. küsimused.
Sõnumite ettevalmistamine:
4. organismide elukeskkond
5.Keskkonnategurid
6. Abiootilised tegurid
7. Biootilised tegurid
8. Antropogeensed tegurid
MBOU Jasnogorski keskkool
Beketova Nurzia Falyakhetdinovna
Bioloogia
10 A klass
Programm algtase haridusasutuste jaoks
Õpik Ponomareva I.N., Kornilova O.A., Loshilina T.E., Izhevsky P.V. Üldine bioloogia
Teema: Elusaine biosfääri organiseerituse taseme tunnused ja roll elu tagamisel Maal.
Sihtmärk: koondab teavet Maa globaalse ökosüsteemi - biosfääri, elusaine organiseerumistaseme biosfääri tunnuste ja rolli kohta elu tagamisel Maal;
Ülesanded:
1. Testida oskust rakendada omandatud teadmisi organisatsiooni biosfääri taseme kohta olukordade põhjendamiseks, oma vaatenurga väljendamiseks ja teaduslikuks põhjendamiseks;
2. Jätkata üldhariduslike oskuste arendamist (tõsta esile peamine, luua põhjus-tagajärg seosed, töötada diagrammidega, teha kindlaks tehtud hinnangute õigsus ning objektide ja nähtuste järjestus);
3. Kujundada tunnetuslikku huvi aine vastu, arendada suhtlemist ja grupitööoskust;
4. Objektiivselt hinnata kooliõpilaste teadmiste ja oskuste taset uuritud rubriigis "Elukorralduse biosfääri tase"
Varustus: tabel “Biosfäär ja selle piirid”, esitlus.
Tundide ajal:
Slaid 1
1. Teadmiste üldistamine ja süstematiseerimine
Vestlus probleemide üle (slaid nr 2)
1. Mis on noosfäär?
2. Kes on noosfääri rajaja?
3. Mis hetkest (teie arvates) hakkas inimene biosfääri (negatiivselt) mõjutama?
4. Mis juhtub, kui ületate biosfääri võimekuse ülemise piiri?
5. tuua näiteid ühiskonna mõjust loodusele, mis tuleb positiivseid kanaleid pidi tagasisidet. Mida sa sellest arvad?
2. Uue materjali õppimine
Tunniplaan:
1. Biosfääri tasandi tunnused.
2. Biosfääri tasandi tunnused.
3. Struktuurielemendid.
4.Põhiprotsessid.
5. Organisatsiooni tunnused.
6. Biosfääri tasandi tähtsus.
3. Konsolideerimine
Õpetaja teeb kokkuvõtte:
Biosfääri elutaset iseloomustavad erilised omadused, keerukusaste ja organiseerituse mustrid, see hõlmab elusorganisme ja nende moodustatavaid looduslikke kooslusi, geograafilist keskkonda ja inimtegevusest tulenevaid tegevusi. Biosfääri tasandil toimuvad väga olulised globaalsed protsessid, mis tagavad elu olemasolu Maal: hapniku moodustumine, päikeseenergia neeldumine ja muundumine, gaasi konstantse koostise säilitamine, biokeemiliste tsüklite ja energiavoo läbiviimine. , liikide ja ökosüsteemide bioloogilise mitmekesisuse areng. Eluvormide mitmekesisus Maal tagab biosfääri stabiilsuse, selle terviklikkuse ja ühtsuse. Elu põhistrateegiaks biosfääri tasandil on elusaine vormide mitmekesisuse ja elu lõpmatuse säilitamine, tagades biosfääri dünaamilise stabiilsuse.
4. Teadmiste kokkuvõte ja jälgimine
Kooliõpilased on oodatud selles rubriigis oma teadmisi ja oskusi proovile panema.
Küsimused
1. Teate, et elusolendite biosfääri organiseerituse tase on kõrgeim ja kõige keerulisem. Loetlege biosfääri tasemele kuuluvad elukorralduse aluseks olevad tasemed nende keerukuse järjekorras.
2. Nimeta märgid, mis võimaldavad iseloomustada biosfääri kui elukorralduse struktuurset taset.
3. Millised on peamised komponendid, mis moodustavad biosfääri struktuuri?
4. Nimeta peamised biosfäärile iseloomulikud protsessid.
5. Miks kuuluvad inimeste majanduslik ja etnokultuuriline tegevus biosfääri peamiste protsesside hulka?
6. Millised nähtused korraldavad biosfääri stabiilsust ehk juhivad selles toimuvaid protsesse?
7. Teadmised sellest, mis on lisaks struktuurile, protsessidele ja korraldusele vajalik biosfääri struktuuri täielikuks mõistmiseks?
8. Sõnasta üldine järeldus Maa elukorralduse biosfääri taseme tähtsusest.
D/z. par.13. küsimused.
Sõnumite ettevalmistamine:
1. inimene kui biosfääri tegur.
2. Biosfääri säilitamise teaduslik alus
3.Säästva arengu väljakutsed
4. organismide elukeskkond
5.Keskkonnategurid
6. Abiootilised tegurid
7. Biootilised tegurid
8. Antropogeensed tegurid
Üldharidusasutuste algtaseme programm
Õpik Ponomareva I.N., Kornilova O.A., Loshilina T.E., Izhevsky P.V. Üldine bioloogia
Teema: Elusaine biosfääri organiseerituse taseme tunnused ja roll elu tagamisel Maal.
Sihtmärk: koondab teavet Maa globaalse ökosüsteemi - biosfääri, elusaine organiseerumistaseme biosfääri tunnuste ja rolli kohta elu tagamisel Maal;
Ülesanded:
1. Testida oskust rakendada omandatud teadmisi organisatsiooni biosfääri taseme kohta olukordade põhjendamiseks, oma vaatenurga väljendamiseks ja teaduslikuks põhjendamiseks;
2. Jätkata üldhariduslike oskuste arendamist (tõsta esile peamine, luua põhjus-tagajärg seosed, töötada diagrammidega, teha kindlaks tehtud hinnangute õigsus ning objektide ja nähtuste järjestus);
3. Kujundada tunnetuslikku huvi aine vastu, arendada suhtlemist ja grupitööoskust;
4. Objektiivselt hinnata kooliõpilaste teadmiste ja oskuste taset uuritud rubriigis "Elukorralduse biosfääri tase"
Varustus: tabel “Biosfäär ja selle piirid”, esitlus.
gi1 kuni Ȯ argin-left:36.0pt;margin-bottom:.0001pt;text-align:justify;text-indent:-18.0pt; line-height:normal;mso-list:l0 level1 lfo1">
Objektiivselt hinnata kooliõpilaste teadmiste ja oskuste taset uuritud rubriigis "Elukorralduse biosfääri tase"
Varustus: tabel “Biosfäär ja selle piirid”, esitlus.
Tundide ajal:
Slaid 1
1. Teadmiste üldistamine ja süstematiseerimine
Vestlus probleemide üle (slaid nr 2)
1. Mis on noosfäär?
2. Kes on noosfääri rajaja?
3. Mis hetkest (teie arvates) hakkas inimene biosfääri (negatiivselt) mõjutama?
4. Mis juhtub, kui ületate biosfääri võimekuse ülemise piiri?
5. Too näiteid ühiskonna mõjust loodusele, mis toimub positiivse tagasiside kanalite kaudu. Mida sa sellest arvad?
2. Uue materjali õppimine
Tunniplaan:
1. Biosfääri tasandi tunnused.
2. Biosfääri tasandi tunnused.
3. Struktuurielemendid.
4.Põhiprotsessid.
5. Organisatsiooni tunnused.
6. Biosfääri tasandi tähtsus.
3. Konsolideerimine
Õpetaja teeb kokkuvõtte:
Biosfääri elutaset iseloomustavad erilised omadused, keerukusaste ja organiseerituse mustrid, see hõlmab elusorganisme ja nende moodustatavaid looduslikke kooslusi, geograafilist keskkonda ja inimtegevusest tulenevaid tegevusi. Biosfääri tasandil toimuvad väga olulised globaalsed protsessid, mis tagavad elu olemasolu Maal: hapniku moodustumine, päikeseenergia neeldumine ja muundumine, gaasi konstantse koostise säilitamine, biokeemiliste tsüklite ja energiavoo läbiviimine. , liikide ja ökosüsteemide bioloogilise mitmekesisuse areng. Eluvormide mitmekesisus Maal tagab biosfääri stabiilsuse, selle terviklikkuse ja ühtsuse. Elu põhistrateegiaks biosfääri tasandil on elusaine vormide mitmekesisuse ja elu lõpmatuse säilitamine, tagades biosfääri dünaamilise stabiilsuse.
4. Teadmiste kokkuvõte ja jälgimine
Kooliõpilased on oodatud selles rubriigis oma teadmisi ja oskusi proovile panema.
Küsimused
1. Teate, et elusolendite biosfääri organiseerituse tase on kõrgeim ja kõige keerulisem. Loetlege biosfääri tasemele kuuluvad elukorralduse aluseks olevad tasemed nende keerukuse järjekorras.
2. Nimeta märgid, mis võimaldavad iseloomustada biosfääri kui elukorralduse struktuurset taset.
3. Millised on peamised komponendid, mis moodustavad biosfääri struktuuri?
4. Nimeta peamised biosfäärile iseloomulikud protsessid.
5. Miks kuuluvad inimeste majanduslik ja etnokultuuriline tegevus biosfääri peamiste protsesside hulka?
6. Millised nähtused korraldavad biosfääri stabiilsust ehk juhivad selles toimuvaid protsesse?
7. Teadmised sellest, mis on lisaks struktuurile, protsessidele ja korraldusele vajalik biosfääri struktuuri täielikuks mõistmiseks?
8. Sõnasta üldine järeldus Maa elukorralduse biosfääri taseme tähtsusest.
D/z. par.13. küsimused.
Sõnumite ettevalmistamine:
1. inimene kui biosfääri tegur.
2. Biosfääri säilitamise teaduslik alus
3.Säästva arengu väljakutsed
4. organismide elukeskkond
5.Keskkonnategurid
6. Abiootilised tegurid
7. Biootilised tegurid
8. Antropogeensed tegurid